
Abstract
Endoplasmic reticulum and nuclear envelope rearrangements after mitosis are often studied in the reconstitution system based on Xenopus egg extract. In our recent work we partially replaced the membrane vesicles in the reconstitution mix with protein-free liposomes to explore the relative contributions of cytosolic and transmembrane proteins. Here we discuss our finding that cytosolic proteins mediate fusion between membranes lacking functional transmembrane proteins and the role of membrane fusion in endoplasmic reticulum and nuclear envelope reorganization. Cytosol-dependent liposome fusion has allowed us to restore, without adding transmembrane nucleoporins, functionality of nuclear pores, their spatial distribution and chromatin decondensation in nuclei formed at insufficient amounts of membrane material and characterized by only partial decondensation of chromatin and lack of nuclear transport. Both the mechanisms and the biological implications of the discovered coupling between spatial distribution of nuclear pores, chromatin decondensation and nuclear transport are discussed.
Acknowledgements
We thank Sergei Pourmal for critical reading of the manuscript. This research was supported by the Intramural Research Program of the Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health (L.V.C.).
Figures and Tables
Figure 1 Mechanism by which peripheral proteins might mediate fusion. (A) Opposing lipid bilayers of biological membranes before fusion. Lipid molecules are depicted as rings (polar groups) with two tails (acyl chains). (B) Fusion proteins (brown-colored shapes with red-colored amphiphilic regions) encircle future fusion site. Shallow insertion of amphiphilic protein regions spreads polar heads of the surrounding lipids. Buckling of the membrane(s) brings membrane bilayers into immediate contact and generates bending stresses at the bulge top(s). (B′) Cross section view from the top. (C) Elastic stresses are released by membrane fusion.
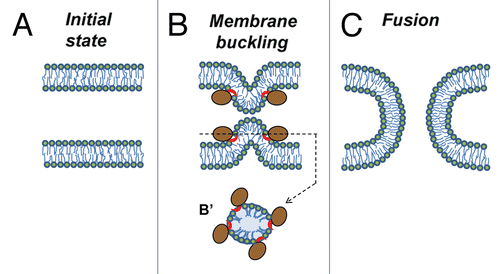
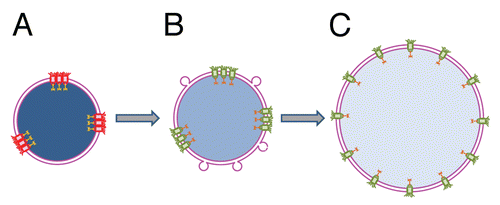
Figure 2 Proposed mechanism of nuclear transport rescue by cytosol-dependent liposome fusion. (A) Shortage of membrane material mechanically restricts chromatin decondensation leading to formation of small undeveloped nuclei enclosed by double membrane NE. Uneven distribution of fully assembled NPCs following uneven distribution of NPC anchoring sites at lamina and chromatin results in functional incompetence of closely spaced pores. (B) Addition of membrane material by cytosol-dependent fusion of liposomes allows additional chromatin decondensation, increases the distances between NPC anchors, and thus, the pores and restores active nuclear transport. (C) Further NE growth is accompanied by full chromatin decondensation and restoration of an even NPC distribution. Functional and non-functional NPCs are shown in green and red, respectively. Degree of chromatin decondensed is depicted by color saturation with less saturated blue color representing more decondensed chromatin.
Extra View to: Rafikova ER, Melikov K, Ramos C, Dye L, Chernomordik LV. Transmembrane protein-free membranes fuse into xenopus nuclear envelope and promote assembly of functional pores. J Biol Chem 2009; 284:29847 - 29859; PMID: 19696024