Abstract
The centrosome is the main microtubule-organizing center in animal cells, and helps to influence the morphology of the microtubule cytoskeleton in interphase and mitosis. The centrosome also templates the assembly of the primary cilium, and together they serve as a nexus of cell signaling that provide cells with diverse organization, motility, and sensory functions. The majority of cells in the human body contain a solitary centrosome and cilium, and cells have evolved regulatory mechanisms to precisely control the numbers of these essential organelles. Defects in the structure and function of cilia lead to a variety of complex disease phenotypes termed ciliopathies, while dysregulation of centrosome number has long been proposed to induce genome instability and tumor formation. Here, we review recent findings that link centrosome amplification to changes in cilium number and signaling capacity, and discuss how supernumerary centrosomes may be an important aspect of a set of cilia-related disease phenotypes.
Keywords: :
Introduction
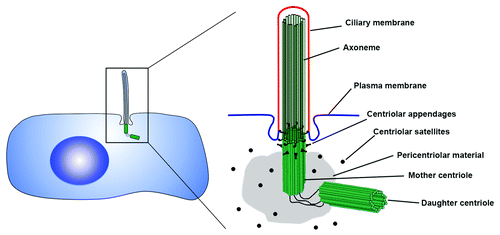
The centrosome is a complex organelle that functions as the major microtubule-organizing center (MTOC) in animal cells. It plays an important structural role in a plethora of cellular processes including intracellular transport, cell polarity, migration and division.Citation1-Citation3 The centrosome also functions as a signaling hub that mediates important aspects of cell physiology, such as orchestrating G1/S transition, entry into mitosis, cytokinesis, and monitoring of DNA damage. At the core of the centrosome is a pair of pinwheel-shaped microtubule structures called centrioles, which are surrounded by pericentriolar material (PCM; ). In animal cells, centrioles contribute to centrosomal functions through recruitment and organization of the PCM, thus facilitating the organization of both the interphase microtubule array as well as the mitotic spindle.Citation4 However, it is becoming increasingly evident that centrioles have evolved for the primary purpose of templating the assembly of cilia,Citation5 essential sensory and motile organelles.
Figure 1. Anatomy of the centrosome-cilium complex. At the core of the centrosome are a pair of centrioles, barrel-shaped structures composed of nine triplet-microtubules. Each centrosome contains a mother and daughter centriole arranged in an orthogonal configuration, which are surrounded by a matrix of proteins called the pericentriolar material (PCM). At the distal end of the older mother centriole are the centriolar appendages, which play important roles in anchoring microtubules as well as tethering the mother centriole to the membrane to induce ciliogenesis. The axoneme of the primary cilium is composed of nine doublet microtubules that grow from the microtubules of the mother centriole, and is surrounded by a ciliary membrane that is distinct from the rest of the plasma membrane.
Except for a few specialized cell types, almost all cells in the human body contain a single centrosome and a single (primary) cilium. The primary cilium is a non-motile microtubule-based organelle that protrudes from the apical surface of most mammalian cells (). Its assembly is templated by the older of the two centrioles in quiescent cells, and it is disassembled as cells re-enter the cell cycle.Citation6-Citation9 Primary cilia are a nexus of cell signaling, associated with regulation of the Notch, Hedgehog, PDGF, TOR and Wnt signaling pathways, among others.Citation10-Citation14 Components of these signaling pathways are concentrated within the ciliary compartment, and this is critical for efficient pathway activation. It is now well established that mutations in proteins that localize to, and function through, cilia cause human disease conditions termed ciliopathies, a group of clinically and genetically overlapping disorders whose etiologies lie in defective ciliary structure and function (for recent reviews seeCitation15-Citation17). Since the assembly of the cilium is dependent on centrioles, mutations that affect the structure and function of centrioles, and the centrosome in general, can also impact normal ciliary function.
Cells control centrosome number using a duplication and segregation mechanism linked to the cell cycle.Citation18,Citation19 Centrosome number is predominantly regulated through the control of centriole duplication. Like DNA, centrioles duplicate in a semi-conservative fashion once and only once per cell cycle, ensuring that each daughter cell inherits one centrosome (with two centrioles) at the end of cell division. The two centrioles present in a quiescent or G1 animal cell differ in age: the older of the two centrioles (which was assembled at least two cell cycles ago) is termed the mother centriole, and the younger centriole (which was assembled in the previous cell cycle) is called the daughter centriole. Centriole duplication begins at the G1 to S phase transition with the formation of one procentriole adjacent to each pre-existing parental centriole.Citation18,Citation20,Citation21 The procentrioles continue to grow during the G2 phase of the cell cycle, and the newly formed centriole pairs move to opposite sides of the cell and help establish the two mitotic spindle poles. During mitosis, the two pairs of centrioles are segregated by the mitotic spindle such that each daughter cell inherits a centriole pair, and thus ends up with one centrosome and the ability to form one primary cilium. To ensure the fidelity of this process, cells have evolved regulatory mechanisms to tightly control the centrosome duplication process throughout the cell cycle.Citation3,Citation18,Citation21 So, what happens when the regulation of centrosome number goes awry?
Centrosome Amplification and Cancer
An important connection between centrosome abnormalities and cancer was first proposed by Theodore Boveri over a century ago.Citation22,Citation23 Since then, researchers have been trying to determine how centrosome defects contribute to cell transformation and tumorigenesis. It has been widely observed that an increase in centrosome number per cell, called centrosome amplification, is a common characteristic of most solid and hematological cancers.Citation24-Citation26 While centrosome amplification is a rarity in healthy adult tissues, the presence of supernumerary centrosomes has been noted in early pre-malignant lesions and correlates with increased tumor grade, size and metastasis in various types of cancers of the brain, breast, colorectum, prostate, kidney, liver and bone.Citation27 These extra centrosomes can arise through several mechanisms: deregulation of the centrosome cycle (resulting in successive rounds of duplication in the same S phase), a failure of cell division (thereby generating a tetraploid cell with two centrosomes), through fusion of two neighboring cells (due to infection by fusogenic viruses), or through unregulated de novo synthesis of centrioles.Citation28-Citation31
The presence of supernumerary centrosomes in tumors raises the question of whether they are simply innocent bystanders or whether they play a causative role in tumor evolution. Extensive work over the past 30 y has attempted to answer this simple yet challenging question, and the findings are summarized in some excellent reviews.Citation24,Citation26,Citation32 Overall, centrosome amplification was proposed to result in the formation of multipolar spindles during mitosis, leading to an increased frequency of aberrant chromosome segregation, causing genetic instability and ultimately driving tumorigenesis. This model persisted, unchallenged, for many years even though it presented a paradox: when multipolar mitosis does occur as a consequence of cells having too many centrosomes, the resulting daughter cells are usually not viable and thus unlikely to contribute to tumor formation.Citation33-Citation36 Indeed, several mechanisms exist that limit the detrimental consequences of supernumerary centrosomes and encourage the formation of a normal bipolar mitosis. These include centrosome removal (extrusion of extra centrosomes from cells), centrosome inactivation (a bipolar mitosis is achieved by silencing MTOC activity of all but 2 centrosomes), and perhaps most importantly centrosome clustering, where cells coalesce their extra centrosomes into two groups to form a bipolar spindle.Citation37-Citation41 So how do supernumerary centrosomes contribute to cell transformation?
Recently, two landmark studies revealed a mechanism by which centrosome amplification may drive tumorigenesis. The authors showed that, even in cells with supernumerary centrosomes that cluster at each pole of the mitotic spindle, the extra centrosomes increase the frequency of merotelic kinetochore attachments, causing chromosome mis-segregation and increased aneuploidy.Citation33,Citation34 Further evidence comes from the demonstration that induction of centrosome amplification can directly initiate tumor formation and metastasis in flies.Citation42 Finally, centrosome amplification has been shown to disrupt cell polarity, migration and cell cycle progression, all hallmarks of transformed cells.Citation32 Thus, it is becoming increasingly evident that rather than serving as a mere beacon of malignancy, supernumerary centrosomes actually drive malignant transformation. However, the recent discovery that centrosome amplification is prevalent in non-cancer related human disease conditions suggests other possible mechanisms by which these supernumerary centrosomes may perturb normal cell function.
Centrosome Amplification in Ciliopathies
Mutations that disrupt the structure and function of cilia are associated with a broad spectrum of human disease phenotypes that include polydactyly, brain malformation, situs inversus (defects of left-right patterning), obesity and polycystic kidney disease.Citation15-Citation17 Due to the critical role of the centrosome as the structural support for primary cilium formation and a coordinator of ciliary protein trafficking, dysregulation of the centrosome often leads to defective ciliary assembly and signaling. Thus, abnormal centrosome function can contribute to cilia-related disease phenotypes. Interestingly, centrosome amplification has recently been reported in tissues from patients and animal models harboring mutations in ciliopathy genes.
Polycystic kidney disease (PKD) is the most common disorder caused by defects in the chemo- and mechano-sensory functions of the primary cilium.Citation43,Citation44 Polycystin-1 (Pkd1) and Polycystin-2 (Pkd2) are membrane-spanning proteins that localize to the centrosome and cilium in renal epithelial cells, and work together in a complex to sense ciliary bending induced by urine flow.Citation45 Mutations in PKD1 and PKD2 cause defects in ciliary signaling, cell polarity, cell division, hyperproliferation of tubular epithelial cells and formation of cysts that profoundly alter the organ architecture and impair renal function.Citation43-Citation45 Recent experiments have uncovered a potentially important role for both Pkd1 and Pkd2 in the maintenance of centrosome integrity. Depletion of Pkd1 was shown to induce centrosome amplification in cells in vitro, while supernumerary centrosomes were observed in kidneys of a Pkd1 knockout animal model and in human renal tissue from Pkd1 patients in vivo.Citation46 The presence of supernumerary centrosomes was noted in seemingly normal tubular cells, suggesting that centrosome amplification is an early event that precedes cyst formation. Similarly, centrosome amplification was reported in fibroblast cells isolated from Pkd2 transgenic mice and in mesenchymal cells of Pkd2 knockout embryos, indicating that Pkd2 dysregulation also alters centrosome number.Citation47
These centrosome number aberrations extend to other cilia-related disease syndromes. Loss of the tumor suppressor genes TSC1 and TSC2, mutations in which cause tuberous sclerosis, also lead to centrosome amplification.Citation48,Citation49 Depletion of the centrosomal Mks1 or Mks3 proteins, mutated in Meckel-Gruber syndrome, similarly results in centrosome amplification.Citation50 Although a causative role for centrosome amplification in the pathogenesis of these disease phenotypes has not been well established, the fact that PKD, TSC and MKS are all attributed to defects in ciliary signaling raises the question of whether changes in centrosome number might affect the structure and/or function of the primary cilia in those cells.
Super-Ciliated Cells
Since it is unclear what phenotypes are conferred upon cells having supernumerary centrosomes that might lead to cilia-related diseases, we considered the possibility that extra centrosomes might result in aberrant cilium number and thus affect normal ciliary signaling. To test this hypothesis, we induced centrosome amplification by briefly expressing the kinase Plk4, which is known to cause formation of multiple centrioles in a diversity of cells and organisms.Citation51-Citation53 As expected, this led to the formation of supernumerary daughter centrioles, which matured into mother centrioles by passage through the cell cycle.Citation54 Remarkably, we discovered that the presence of extra centrosomes invariably led to the formation of more than one primary cilium per cell (). These super-ciliated cells contained up to six primary cilia per cell.Citation54
Figure 2. Supernumerary centrosomes nucleate extra cilia and compromise ciliary signaling. (A) Examples of fibroblast cells with amplified centrioles and cilia. Centriole amplification was achieved by briefly expressing Plk4.Citation54 Cells assembled excess centrioles in S-phase, which clustered together at the poles of the mitotic spindle. Once cells completed mitosis and entered a quiescent stage, the majority of cells possessed more than one primary cilium. Cells were stained for glutamylated tubulin (centrioles and cilia; red) and DNA (blue). Images were reproduced with permission from reference 54. (B) Model depicting ciliary dysfunction caused by dilution of ciliary signaling component (red) in super-ciliated cells. In normal mono-ciliated cells, ciliary proteins are concentrated within the ciliary compartment, and this is critical for pathway activation. In super-ciliated cells, the ciliary protein is distributed among multiple cilia, resulting in a reduced concentration of the signaling component per unit length cilium, causing defective pathway activation.Citation54
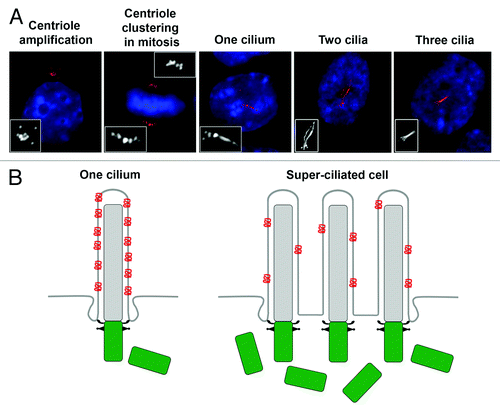
Next, we wondered whether an increase in the number of primary cilia might perturb signaling pathways that rely on cilium function. We quantified the concentration of signaling molecules that reside within the primary cilium, focusing first on the transmembrane protein Smoothened (Smo). In response to Sonic Hedgehog (Shh) ligand, Smo translocates into the cilium,Citation55 and this is essential in activating downstream signaling in mammalian cells.Citation56 Using quantitative single-cell immunofluorescence microscopy, we determined that the amount of Smo per unit length cilium decreased proportionally as the number of cilia per cell increased.Citation54 We refer to this decrease in ciliary protein concentration as a “ciliary dilution” phenotype, and in the case of Smo, this dilution resulted in defective activation of the Shh pathway.Citation54
The observation that supernumerary centrosomes resulted in the formation of extra cilia and caused defective recruitment of ciliary Smo protein was intriguing, and raised the question of whether other signaling pathways are similarly affected. To address the generality and nature of the ciliary dilution phenotype, we examined the ciliary concentrations of additional proteins that localize to cilia in super-ciliated cells. We found that the serotonin 6 (Htr6) receptor,Citation57,Citation58 the fibrocystin (PKHD) protein,Citation59 and the GTPase Arl13bCitation60 all exhibited a ciliary dilution phenotype similar to that of Smo in super-ciliated cells.Citation54 The ciliary dilution phenotype was observed even under conditions in which the total concentration of the transported protein was not limiting. Because super-ciliated cells assembled cilia of similar length to those in mono-ciliated cells, we reasoned that components of the ciliary machinery might not display the ciliary dilution phenotype. Consistent with this hypothesis, the ciliary concentration of IFT88, a component of the intraflagellar transport machinery required for cilium assembly,Citation61 was equal in mono- and super-ciliated cells. Lastly, we found that the ciliary dilution phenotype was dependent on number of cilia rather than number of centrioles, suggesting that the mechanism determining ciliary protein levels assesses the number of cilia per cell, rather than the number of centrosomes per cell.Citation54
Our experiments demonstrate that centrosome amplification can cause the formation of ectopic cilia, and leads to a reduction in ciliary signaling capacity. To determine the functional consequences of having too many cilia in a cell, we tested the effect of extra cilia on cellular processes that depend on cilium function. Ciliary signaling is essential in organizing the architecture and polarity of epithelial cells through regulation of both the canonical and non-canonical (planar cell polarity) Wnt signaling pathways.Citation62 We found that the presence of supernumerary centrosomes and cilia disrupted epithelial cell polarity and organization in vitro.Citation54 Furthermore, super-ciliated cells displayed slow progression through the cell cycle as well as defective directional cell migration (our unpublished results), both processes that rely on proper ciliary function.Citation11,Citation63,Citation64 It is important to note that the observed deficiencies in cell polarity, migration and division in super-ciliated cells are similar to cellular defects caused by mutations in canonical ciliopathy genes. It is tempting to speculate that the reduction in ciliary protein concentration, due to supernumerary centrosomes and cilia, might lead to the same phenotypic outcome in patients.
Concluding Remarks
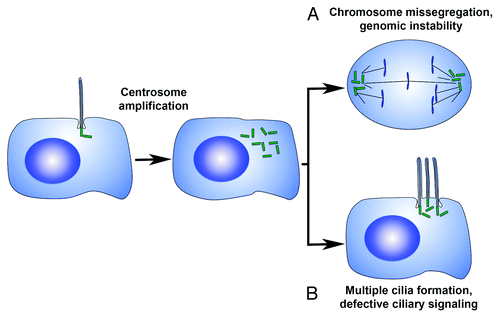
These results suggest a mechanism by which the presence of supernumerary centrosomes may contribute to defective ciliary signaling, possibly through formation of ectopic primary cilia (). Recent studies have reported findings that support this theory. That the presence of extra cilia can lead to altered ciliary signaling capacity is reinforced by results demonstrating a reduction in canonical Wnt pathway activation in super-ciliated cells.Citation65 Importantly, the presence of super-ciliated cells due to centrosome amplification has now been reported in multiple tissues and cell types.
Figure 3. Mechanisms by which centrosome amplification may contribute to cell transformation. (A) Supernumerary centrosomes can lead to an increase in the frequency of lagging chromosomes, inducing aneuploidy through chromosome mis-segregation. (B) Centrosome amplification can modulate the number of primary cilia per cell, resulting in compromised ciliary signaling.
Kidney tissue from patients bearing a mutation in MKS3 were found to contain cells with extra centrioles and cilia in vivo,Citation50 and kidney cysts in patients with mutations in PKD1 display gross centrosome amplification and contain super-ciliated cells in vivo (our unpublished results). Mutations in TSC1 and TSC2 result in enhanced ciliary formation, and cells isolated from TSC2−/− mice have supernumerary centrosomes, form extra cilia and display the same ciliary protein dilution phenotype.Citation48,Citation49,Citation54 Mutations in the ciliary protein Inversin cause nephronophthisis, an autosomal recessive form of polycystic kidney disease.Citation66 Deletion of Inv in the developing mouse nephron leads to defective cytokinesis and an increased frequency of binucleate cells that contain extra cilia in vivo.Citation67 Similarly, loss of Citron kinase in rats, which causes failed cytokinesis of cortical neural progenitor cells, results in the formation of binucleate neurons that also possess supernumerary centrioles and multiple primary cilia per neuron.Citation68 The authors speculate that the presence of multiple cilia in the multinucleated neurons may compromise the ability of those cells to respond to extracellular stimuli.Citation68 In line with these results, we found that blocking cytokinesis in fibroblast and epithelial cells also yielded binucleate cells, formed two cilia per cell and displayed the same dilution of ciliary signaling molecules.Citation54
Together, these studies indicate that regardless of the mechanism leading to centrosome amplification, cells with excess centrosomes often form more than one primary cilium. This phenomenon has been observed in various cell types and tissues, both in vitro and in vivo. It raises the interesting possibility that aberrant primary cilium-associated signaling, due to the presence of ectopic centrosomes and cilia, might be an important aspect of a set of cilia-related disease phenotypes. However, a causal role for centrosome amplification in cystogenesis, or other ciliopathy phenotypes, has not been established. Similarly, it is not known whether the presence of excess cilia is in fact contributing to cell transformation, cyst formation or tumorigenesis in vivo. Future work, particularly at the organism level, will be essential to ascertain the impact of centrosome amplification on ciliary function during development, and how it may contribute to disease.
Questions and Answers
Dr Helen Liapis, Professor of Pathology and Immunology, Washington University School of Medicine
Moe this was a wonderful talk. Beautiful pictures, I am using the last one you showed in the new Heptinstall 's Pathology of the Kidney textbook; it is probably going to be on the cover. I still don’t understand why there are no data in human disease, particularly polycystic kidney disease, to show structural abnormalities of the cilia. As you know, this has cast doubts of the relevance of ciliary biology (which is beautiful) to human disease. Why do you think that is?
Dr Mahjoub
Actually, structural defects in cilia have been observed in a number of cilia-related diseases. If you were to completely abrogate cilia structure, you get embryonic lethality, so there is a reason why there aren’t humans walking around completely lacking cilia. But there are specific sub domains within the cilium, like the transition zone, the periciliary necklace, and the ciliary pocket that control trafficking of proteins in and out of the cilium. These are structural components that you may not see by EM per se, but there are proteins that reside in, and help define, these unique regions. These proteins are commonly mutated in ciliopathies, and we can count those as structural defects. So even though the cilium in itself is fully assembled, the defects in those little structures change which proteins get in and change which proteins go out, and that alters the activity of the signaling pathways downstream of those proteins. So you might not think of them as a structural defect in the gross term but they still impact the signaling capacity of the cilium.
Dr Ying Chen, Assistant Professor Medicine, Washington University School of Medicine
So Moe, I’m thinking that in autosomal dominant polycystic kidney disease, research has proven that there is a phenotypic switch in the second messenger of cAMP, which alters the state of cells from the non-proliferative to pro-proliferative state. Have you checked your multi-ciliated cells to see if there is an impact in cAMP signaling?
Dr Mahjoub
Although we have not examined cAMP signaling, we did look at cell proliferation and noticed a difference. Cells that have multiple primary cilia divide less often than cells that have one cilium. Our original hypothesis was that the presence of more cilia in a cell would lead to increased primary cilium signaling, and potentially more cell division. But what we found is that more cilia actually led to less signaling, and less cell division. But I only showed you data for pro-proliferative pathways. If we consider inhibitory pathways that function through cilia, then the presence of supernumerary cilia would presumably dilute those proteins in the same way, so you could have a release of inhibition on cell proliferation. But we have not tested that yet.
Dr Jeffrey Miner, Professor of Medicine, Washington University School of Medicine
Can you bypass the dampened sonic hedgehog signaling when you have the abnormal multiple cilia by overexpressing Smoothened?
Dr Mahjoub
Although we did not test it specifically for the Shh pathway, overexpression of ciliary membrane proteins does not lead to more of those proteins localizing in cilia. We did overexpress a number of ciliary signaling molecules, and although you can have lots of them in the cytoplasm, they do not get imported into the cilium. At first we thought that one of the trafficking components might become saturated, such that you can have all the cargo you want in the cell body, but only so much of it could be loaded onto the trafficking machinery. That doesn’t seem to be the case. The experiments in which we unclustered the centrioles, so that the multiple cilia in the cell were separated and residing in their own ciliary pocket proved that point. In those cells, proteins were targeted normally, so the limiting factor was the structure of the ciliary pocket itself. Think of it as a little train station: normally, cargo only has one train (cilium) to get loaded onto, but in super-ciliated cells there are three or more trains for the cargo to get loaded on, so it gets distributed differentially among the multiple trains (cilia).
Dr Keith Hruska, Professor of Pediatrics and Medicine, Washington University School of Medicine
One clinical insight into ADPKD comes from the skeleton, where the osteocyte cilium is responsible for transmitting mechanical load. What the clinical part is, we find that there is a tremendous decrease in remodeling rate in the ADPKD osteocytes, both in vivo and in vitro, and related to that there is an increase in FGF23 secretion. So when you are talking about Smoothened in the signaling pathways, here the signaling pathway is Wnt since it is the main anabolic factor in the skeleton. So have you had an opportunity to look at Wnt pathways?
Dr Mahjoub
We did not look directly at Wnt pathway activity in the super-ciliated cells, although we did the functional assay where we showed that cell polarity (canonical and planar cell polarity) was disrupted in those cells. We picked Sonic Hedgehog because it is very well characterized with regards to its role in the cilium. We used that as a tool to test the activity of that pathway and then we looked at a number of other ciliary signaling molecules just to show that it is likely happening to other pathways too. I should mention that a recent paper from the Gleeson lab demonstrated a reduction in canonical Wnt pathway activation in super-ciliated cells,Citation65 which nicely supports our findings.
Dr Feng Chen, Associate Professor of Medicine, Washington University School of Medicine
A couple of questions. The first one is really simple but I am curious. You mentioned that during asymmetric divisions in mono-ciliated cells, the older mother centriole is always inherited by one of the daughter cells, the stem cell for example. I wonder how the centrioles are distributed within the super-ciliated cells. Would all the older centrioles and all of the cilia within a cell go to one daughter cell when the cell divides?
Dr Mahjoub
The asymmetric inheritance of centrioles based on their age has been reported only in certain cell types so far, such as neural progenitor cells. However, this process is not occurring in our cultured cells in vitro, so I cannot address that directly. What happens in our experiments is that we induce the formation of multiple daughter centrioles at once, which are template by the original mother and original daughter centrioles. Instead of the normal duplication cycle, where you get 1-to-1 ratios of new daughter centrioles to parental centrioles, these now make 5 or 6 new daughter centrioles each so you got a bunch of young centrioles at once. The cells then go through mitosis at least twice, and a fraction of them become mother centrioles because they have undergone maturation. An interesting experiment would be to induce centriole amplification in certain cell types (like neural progenitors), and see whether the very oldest mother centriole and the original daughter centriole are segregated differentially, even if extra centrioles are around. We would have to do it in a cell type that has asymmetric cell division as part of its program, and see if that disrupts cell fate. The cells we are looking at do not divide this way, they are just epithelial or fibroblast cells that divide symmetrically.
Dr Marc Hammerman, Chromalloy Professor, Washington University School of Medicine
How many generations of centrioles are there? Is there such a thing as a great grandmother centriole?
Dr Mahjoub
In theory, we can trace the lineage of centrioles in dividing cells through time indefinitely, although it is technically challenging. A cell starts out with a mother and a daughter centriole, builds a pair of new daughter centrioles in S-phase then segregates the two pairs to its sister cells following division. This means that the original mother centriole will always be the oldest centriole in the entire generation of cells. In theory, one should be able to follow the location of that centriole forever because every time that cell divides, one of the two cells will inherit that oldest centriole, because these organelles do not disassemble and reassemble. We should be able to trace their lineage. What that means to the biology of that cell, we do not know yet.
Hani Suleiman, Staff Scientist, Department of Pathology and Immunology Washington University School of Medicine
Regarding the directional migration, could you elaborate on why you would have directional migration defects if you still have the 3 or 2 cilia in the same spot at the leading edge of a polarized cell?
Dr Mahjoub
Dr. Soren Christensen’s lab has done a lot of good work to show that the cilium responds to PDGF ligand through the PDGF-α receptor, which is localized to the cilium.Citation63 So the primary cilium appears to be important for receiving extracellular signals that help orient cells during migration, for example during would healing. I showed you that having extra cilia causes defects in the signaling capacity of those cilia, because there is less of each signaling protein, and thus less interaction among the components. Therefore, the cell migration experiments were another way to test the functionality of the cilia in super-ciliated cells. Presumably, these cilia have less PDGF receptor per unit length just like the other proteins we looked at, and are not able to receive that extracellular signal very well. The result is that cells do not migrate properly; although they fill in the wound over time in our scratch assay, the directionality of migration is perturbed. One experiment we would like to do is to examine this at the single cell level, instead of the tissue level. We would feed the cells a specific ligand (e.g., PDGF) and track its movement toward or away from the stimulus. We would do this with cells that have one or more cilia and see how each cell responds.
| Abbreviations: | ||
| MTOC | = | microtubule-organizing center |
| PCM | = | pericentriolar material |
| PKD | = | polycystic kidney disease |
| TSC | = | tuberous sclerosis |
| MKS | = | Meckel-Gruber syndrome |
| Shh | = | Sonic hedgehog |
| Smo | = | smoothened |
Acknowledgments
I wish to thank Tim Stearns (Stanford University) and members of my lab for critical comments on the manuscript, and apologize to colleagues whose work was not discussed or cited owing to space constraints.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Note
Edited transcripts of research conferences sponsored by Organogenesis and the Washington University George M. O’Brien Center for Kidney Disease Research (P30 DK079333) are published in Organogenesis. These conferences cover organogenesis in all multicellular organisms including research into tissue engineering, artificial organs and organ substitutes and are participated in by faculty at Washington University School of Medicine, St. Louis Missouri USA.
References
- Bornens M. Centrosome composition and microtubule anchoring mechanisms. Curr Opin Cell Biol 2002; 14:25 - 34; http://dx.doi.org/10.1016/S0955-0674(01)00290-3; PMID: 11792541
- Lüders J, Stearns T. Microtubule-organizing centres: a re-evaluation. Nat Rev Mol Cell Biol 2007; 8:161 - 7; http://dx.doi.org/10.1038/nrm2100; PMID: 17245416
- Nigg EA, Stearns T. The centrosome cycle: Centriole biogenesis, duplication and inherent asymmetries. Nat Cell Biol 2011; 13:1154 - 60; http://dx.doi.org/10.1038/ncb2345; PMID: 21968988
- Gönczy P. Towards a molecular architecture of centriole assembly. Nat Rev Mol Cell Biol 2012; 13:425 - 35; http://dx.doi.org/10.1038/nrm3373; PMID: 22691849
- Debec A, Sullivan W, Bettencourt-Dias M. Centrioles: active players or passengers during mitosis?. Cell Mol Life Sci 2010; 67:2173 - 94; http://dx.doi.org/10.1007/s00018-010-0323-9; PMID: 20300952
- Garcia-Gonzalo FR, Reiter JF. Scoring a backstage pass: mechanisms of ciliogenesis and ciliary access. J Cell Biol 2012; 197:697 - 709; http://dx.doi.org/10.1083/jcb.201111146; PMID: 22689651
- Tucker RW, Pardee AB, Fujiwara K. Centriole ciliation is related to quiescence and DNA synthesis in 3T3 cells. Cell 1979; 17:527 - 35; http://dx.doi.org/10.1016/0092-8674(79)90261-7; PMID: 476831
- Ishikawa H, Marshall WF. Ciliogenesis: building the cell’s antenna. Nat Rev Mol Cell Biol 2011; 12:222 - 34; http://dx.doi.org/10.1038/nrm3085; PMID: 21427764
- Seeley ES, Nachury MV. The perennial organelle: assembly and disassembly of the primary cilium. J Cell Sci 2010; 123:511 - 8; http://dx.doi.org/10.1242/jcs.061093; PMID: 20144999
- Goetz SC, Anderson KV. The primary cilium: a signalling centre during vertebrate development. Nat Rev Genet 2010; 11:331 - 44; http://dx.doi.org/10.1038/nrg2774; PMID: 20395968
- Schneider L, Clement CA, Teilmann SC, Pazour GJ, Hoffmann EK, Satir P, et al. PDGFRalphaalpha signaling is regulated through the primary cilium in fibroblasts. Curr Biol 2005; 15:1861 - 6; http://dx.doi.org/10.1016/j.cub.2005.09.012; PMID: 16243034
- Simons M, Gloy J, Ganner A, Bullerkotte A, Bashkurov M, Krönig C, et al. Inversin, the gene product mutated in nephronophthisis type II, functions as a molecular switch between Wnt signaling pathways. Nat Genet 2005; 37:537 - 43; http://dx.doi.org/10.1038/ng1552; PMID: 15852005
- Habbig S, Bartram MP, Müller RU, Schwarz R, Andriopoulos N, Chen S, et al. NPHP4, a cilia-associated protein, negatively regulates the Hippo pathway. J Cell Biol 2011; 193:633 - 42; http://dx.doi.org/10.1083/jcb.201009069; PMID: 21555462
- Ezratty EJ, Stokes N, Chai S, Shah AS, Williams SE, Fuchs E. A role for the primary cilium in Notch signaling and epidermal differentiation during skin development. Cell 2011; 145:1129 - 41; http://dx.doi.org/10.1016/j.cell.2011.05.030; PMID: 21703454
- Gerdes JM, Davis EE, Katsanis N. The vertebrate primary cilium in development, homeostasis, and disease. Cell 2009; 137:32 - 45; http://dx.doi.org/10.1016/j.cell.2009.03.023; PMID: 19345185
- Nigg EA, Raff JW. Centrioles, centrosomes, and cilia in health and disease. Cell 2009; 139:663 - 78; http://dx.doi.org/10.1016/j.cell.2009.10.036; PMID: 19914163
- Waters AM, Beales PL. Ciliopathies: an expanding disease spectrum. Pediatr Nephrol 2011; 26:1039 - 56; http://dx.doi.org/10.1007/s00467-010-1731-7; PMID: 21210154
- Azimzadeh J, Marshall WF. Building the centriole. Curr Biol 2010; 20:R816 - 25; http://dx.doi.org/10.1016/j.cub.2010.08.010; PMID: 20869612
- Hatch E, Stearns T. The life cycle of centrioles. Cold Spring Harb Symp Quant Biol 2010; 75:425 - 31; http://dx.doi.org/10.1101/sqb.2010.75.054; PMID: 21502410
- Pelletier L, Yamashita YM. Centrosome asymmetry and inheritance during animal development. Curr Opin Cell Biol 2012; 24:541 - 6; http://dx.doi.org/10.1016/j.ceb.2012.05.005; PMID: 22683192
- Brito DA, Gouveia SM, Bettencourt-Dias M. Deconstructing the centriole: structure and number control. Curr Opin Cell Biol 2012; 24:4 - 13; http://dx.doi.org/10.1016/j.ceb.2012.01.003; PMID: 22321829
- Boveri T. Ueber mehrpolige Mitosen als Mittel zur Analyse des Zellkerns. Verh Phys-med Ges Wurzburg NF 1902; 35:67 - 90
- Boveri T. Concerning the origin of malignant tumours by Theodor Boveri. Translated and annotated by Henry Harris. J Cell Sci 2008; 121:Suppl 1 1 - 84; http://dx.doi.org/10.1242/jcs.025742; PMID: 18089652
- Godinho SA, Kwon M, Pellman D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Cancer Metastasis Rev 2009; 28:85 - 98; http://dx.doi.org/10.1007/s10555-008-9163-6; PMID: 19156503
- Krämer A, Neben K, Ho AD. Centrosome aberrations in hematological malignancies. Cell Biol Int 2005; 29:375 - 83; http://dx.doi.org/10.1016/j.cellbi.2005.03.004; PMID: 15996491
- Marthiens V, Piel M, Basto R. Never tear us apart--the importance of centrosome clustering. J Cell Sci 2012; 125:3281 - 92; http://dx.doi.org/10.1242/jcs.094797; PMID: 22956721
- Chan JY. A clinical overview of centrosome amplification in human cancers. Int J Biol Sci 2011; 7:1122 - 44; http://dx.doi.org/10.7150/ijbs.7.1122; PMID: 22043171
- Duelli D, Lazebnik Y. Cell-to-cell fusion as a link between viruses and cancer. Nat Rev Cancer 2007; 7:968 - 76; http://dx.doi.org/10.1038/nrc2272; PMID: 18034186
- Fujiwara T, Bandi M, Nitta M, Ivanova EV, Bronson RT, Pellman D. Cytokinesis failure generating tetraploids promotes tumorigenesis in p53-null cells. Nature 2005; 437:1043 - 7; http://dx.doi.org/10.1038/nature04217; PMID: 16222300
- Storchova Z, Pellman D. From polyploidy to aneuploidy, genome instability and cancer. Nat Rev Mol Cell Biol 2004; 5:45 - 54; http://dx.doi.org/10.1038/nrm1276; PMID: 14708009
- Nigg EA. Centrosome aberrations: cause or consequence of cancer progression?. Nat Rev Cancer 2002; 2:815 - 25; http://dx.doi.org/10.1038/nrc924; PMID: 12415252
- Ogden A, Rida PC, Aneja R. Heading off with the herd: how cancer cells might maneuver supernumerary centrosomes for directional migration. Cancer Metastasis Rev 2013; 32:269 - 87; http://dx.doi.org/10.1007/s10555-012-9413-5; PMID: 23114845
- Ganem NJ, Godinho SA, Pellman D. A mechanism linking extra centrosomes to chromosomal instability. Nature 2009; 460:278 - 82; http://dx.doi.org/10.1038/nature08136; PMID: 19506557
- Silkworth WT, Nardi IK, Scholl LM, Cimini D. Multipolar spindle pole coalescence is a major source of kinetochore mis-attachment and chromosome mis-segregation in cancer cells. PLoS One 2009; 4:e6564; http://dx.doi.org/10.1371/journal.pone.0006564; PMID: 19668340
- Weaver BA, Cleveland DW. Aneuploidy: instigator and inhibitor of tumorigenesis. Cancer Res 2007; 67:10103 - 5; http://dx.doi.org/10.1158/0008-5472.CAN-07-2266; PMID: 17974949
- Weaver BA, Silk AD, Montagna C, Verdier-Pinard P, Cleveland DW. Aneuploidy acts both oncogenically and as a tumor suppressor. Cancer Cell 2007; 11:25 - 36; http://dx.doi.org/10.1016/j.ccr.2006.12.003; PMID: 17189716
- Gräf R, Euteneuer U, Ho TH, Rehberg M. Regulated expression of the centrosomal protein DdCP224 affects microtubule dynamics and reveals mechanisms for the control of supernumerary centrosome number. Mol Biol Cell 2003; 14:4067 - 74; http://dx.doi.org/10.1091/mbc.E03-04-0242; PMID: 14517319
- Iwao Y, Murakawa T, Yamaguchi J, Yamashita M. Localization of gamma-tubulin and cyclin B during early cleavage in physiologically polyspermic newt eggs. Dev Growth Differ 2002; 44:489 - 99; http://dx.doi.org/10.1046/j.1440-169X.2002.00661.x; PMID: 12492507
- Brinkley BR. Managing the centrosome numbers game: from chaos to stability in cancer cell division. Trends Cell Biol 2001; 11:18 - 21; http://dx.doi.org/10.1016/S0962-8924(00)01872-9; PMID: 11146294
- Ring D, Hubble R, Kirschner M. Mitosis in a cell with multiple centrioles. J Cell Biol 1982; 94:549 - 56; http://dx.doi.org/10.1083/jcb.94.3.549; PMID: 7130271
- Kwon M, Godinho SA, Chandhok NS, Ganem NJ, Azioune A, Thery M, et al. Mechanisms to suppress multipolar divisions in cancer cells with extra centrosomes. Genes Dev 2008; 22:2189 - 203; http://dx.doi.org/10.1101/gad.1700908; PMID: 18662975
- Basto R, Brunk K, Vinadogrova T, Peel N, Franz A, Khodjakov A, et al. Centrosome amplification can initiate tumorigenesis in flies. Cell 2008; 133:1032 - 42; http://dx.doi.org/10.1016/j.cell.2008.05.039; PMID: 18555779
- Winyard P, Jenkins D. Putative roles of cilia in polycystic kidney disease. Biochim Biophys Acta 2011; 1812:1256 - 62; http://dx.doi.org/10.1016/j.bbadis.2011.04.012; PMID: 21586324
- Kotsis F, Boehlke C, Kuehn EW. The ciliary flow sensor and polycystic kidney disease. Nephrol Dial Transplant 2013; 28:518 - 26; http://dx.doi.org/10.1093/ndt/gfs524; PMID: 23314319
- Nauli SM, Alenghat FJ, Luo Y, Williams E, Vassilev P, Li X, et al. Polycystins 1 and 2 mediate mechanosensation in the primary cilium of kidney cells. Nat Genet 2003; 33:129 - 37; http://dx.doi.org/10.1038/ng1076; PMID: 12514735
- Battini L, Macip S, Fedorova E, Dikman S, Somlo S, Montagna C, et al. Loss of polycystin-1 causes centrosome amplification and genomic instability. Hum Mol Genet 2008; 17:2819 - 33; http://dx.doi.org/10.1093/hmg/ddn180; PMID: 18566106
- Burtey S, Riera M, Ribe E, Pennenkamp P, Rance R, Luciani J, et al. Centrosome overduplication and mitotic instability in PKD2 transgenic lines. Cell Biol Int 2008; 32:1193 - 8; http://dx.doi.org/10.1016/j.cellbi.2008.07.021; PMID: 18725310
- Astrinidis A, Senapedis W, Henske EP. Hamartin, the tuberous sclerosis complex 1 gene product, interacts with polo-like kinase 1 in a phosphorylation-dependent manner. Hum Mol Genet 2006; 15:287 - 97; http://dx.doi.org/10.1093/hmg/ddi444; PMID: 16339216
- Hartman TR, Liu D, Zilfou JT, Robb V, Morrison T, Watnick T, et al. The tuberous sclerosis proteins regulate formation of the primary cilium via a rapamycin-insensitive and polycystin 1-independent pathway. Hum Mol Genet 2009; 18:151 - 63; http://dx.doi.org/10.1093/hmg/ddn325; PMID: 18845692
- Tammachote R, Hommerding CJ, Sinders RM, Miller CA, Czarnecki PG, Leightner AC, et al. Ciliary and centrosomal defects associated with mutation and depletion of the Meckel syndrome genes MKS1 and MKS3. Hum Mol Genet 2009; 18:3311 - 23; http://dx.doi.org/10.1093/hmg/ddp272; PMID: 19515853
- Habedanck R, Stierhof Y-D, Wilkinson CJ, Nigg EA. The Polo kinase Plk4 functions in centriole duplication. Nat Cell Biol 2005; 7:1140 - 6; http://dx.doi.org/10.1038/ncb1320; PMID: 16244668
- Pearson CG, Winey M. Plk4/SAK/ZYG-1 in the regulation of centriole duplication. F1000 Biol Rep 2010; 2:58; PMID: 21173875
- Sillibourne JE, Bornens M. Polo-like kinase 4: the odd one out of the family. Cell Div 2010; 5:25; http://dx.doi.org/10.1186/1747-1028-5-25; PMID: 20920249
- Mahjoub MR, Stearns T. Supernumerary centrosomes nucleate extra cilia and compromise primary cilium signaling. Curr Biol 2012; 22:1628 - 34; http://dx.doi.org/10.1016/j.cub.2012.06.057; PMID: 22840514
- Corbit KC, Aanstad P, Singla V, Norman AR, Stainier DYR, Reiter JF. Vertebrate Smoothened functions at the primary cilium. Nature 2005; 437:1018 - 21; http://dx.doi.org/10.1038/nature04117; PMID: 16136078
- Rohatgi R, Milenkovic L, Scott MP. Patched1 regulates hedgehog signaling at the primary cilium. Science 2007; 317:372 - 6; http://dx.doi.org/10.1126/science.1139740; PMID: 17641202
- Hamon M, Doucet E, Lefèvre K, Miquel MC, Lanfumey L, Insausti R, et al. Antibodies and antisense oligonucleotide for probing the distribution and putative functions of central 5-HT6 receptors. Neuropsychopharmacology 1999; 21:Suppl 68S - 76S; PMID: 10432491
- Berbari NF, Johnson AD, Lewis JS, Askwith CC, Mykytyn K. Identification of ciliary localization sequences within the third intracellular loop of G protein-coupled receptors. Mol Biol Cell 2008; 19:1540 - 7; http://dx.doi.org/10.1091/mbc.E07-09-0942; PMID: 18256283
- Follit JA, Li L, Vucica Y, Pazour GJ. The cytoplasmic tail of fibrocystin contains a ciliary targeting sequence. J Cell Biol 2010; 188:21 - 8; http://dx.doi.org/10.1083/jcb.200910096; PMID: 20048263
- Lim YS, Chua CE, Tang BL. Rabs and other small GTPases in ciliary transport. Biol Cell 2011; 103:209 - 21; http://dx.doi.org/10.1042/BC20100150; PMID: 21488838
- Pazour GJ, Dickert BL, Vucica Y, Seeley ES, Rosenbaum JL, Witman GB, et al. Chlamydomonas IFT88 and its mouse homologue, polycystic kidney disease gene tg737, are required for assembly of cilia and flagella. J Cell Biol 2000; 151:709 - 18; http://dx.doi.org/10.1083/jcb.151.3.709; PMID: 11062270
- Wallingford JB, Mitchell B. Strange as it may seem: the many links between Wnt signaling, planar cell polarity, and cilia. Genes Dev 2011; 25:201 - 13; http://dx.doi.org/10.1101/gad.2008011; PMID: 21289065
- Christensen ST, Pedersen SF, Satir P, Veland IR, Schneider L. The primary cilium coordinates signaling pathways in cell cycle control and migration during development and tissue repair. Curr Top Dev Biol 2008; 85:261 - 301; http://dx.doi.org/10.1016/S0070-2153(08)00810-7; PMID: 19147009
- Goto H, Inoko A, Inagaki M. Cell cycle progression by the repression of primary cilia formation in proliferating cells. Cell Mol Life Sci 2013; http://dx.doi.org/10.1007/s00018-013-1302-8; PMID: 23475109
- Lancaster MA, Schroth J, Gleeson JG. Subcellular spatial regulation of canonical Wnt signalling at the primary cilium. Nat Cell Biol 2011; 13:700 - 7; http://dx.doi.org/10.1038/ncb2259; PMID: 21602792
- Lienkamp S, Ganner A, Walz G. Inversin, Wnt signaling and primary cilia. Differentiation 2012; 83:S49 - 55; http://dx.doi.org/10.1016/j.diff.2011.11.012; PMID: 22206729
- Werner M, Ward HH, Phillips CL, Miller C, Gattone VH 2nd, Bacallao RL. Inversin modulates the cortical actin network during mitosis. Am J Physiol Cell Physiol 2013; http://dx.doi.org/10.1152/ajpcell.00279.2012; PMID: 23515530
- Anastas SB, Mueller D, Semple-Rowland SL, Breunig JJ, Sarkisian MR. Failed cytokinesis of neural progenitors in citron kinase-deficient rats leads to multiciliated neurons. Cereb Cortex 2011; 21:338 - 44; http://dx.doi.org/10.1093/cercor/bhq099; PMID: 20525772