Abstract
Secreted signaling molecules of the Wnt family have been found to play a central role in controlling embryonic development of a wide range of taxa from Hydra to humans. The most extensively studied Wnt signaling pathway is the canonical Wnt pathway, which controls gene expression by stabilizing β-catenin, and regulates a multitude of developmental processes. More recently, noncanonical Wnt pathways, which are β-catenin-independent, have been found to be important developmental regulators. Understanding the mechanisms of Wnt signaling is essential for the development of novel preventive and therapeutic approaches of human diseases. Limb development is a paradigm to study the principles of Wnt signaling in various developmental contexts. In the developing vertebrate limb, Wnt signaling has been shown to have important functions during limb bud initiation, limb outgrowth, early limb patterning, and later limb morphogenesis events. This review provides a brief overview on the diversity of Wnt-dependent signaling events during embryonic development of the vertebrate limb.
Acknowledgments
Work in our laboratory is sponsored by the Deutsche Forschungsgemeinschaft DFG (SFB592, GRK1104 to M.S.) and the European NoE MYORES (M.S.).
Figures and Tables
Fig 1 Three Wnt signaling pathways (modified from Huelsken and Behrens, 2002). (A) The Wnt/β-catenin pathway. Wnt signal is transduced through the frizzled receptor (Fz) via dishevelled (Dsh), which represses the axin/glycogen synthase kinase-3β (GSK3β) complex, which induces the degradation of β-catenin. Accumulated Cytoplasmic β-catenin is translocated to the nucleus where it binds with Tcf/Lef, thus activating the transcription of its target gene. (B) The planar cell polarity (PCP) pathway activated in response to Wnt signaling signals via the small GTPases Rho and Cdc42 to c-Jun N-terminal kinase (JNK), acting primarily on the cytoskeleton (C) An alternative pathway stimulates the release of intracellular Ca2+, activating protein kinase C (PKC) and Ca2+/calmodulin-dependent kinase II (CamKII).
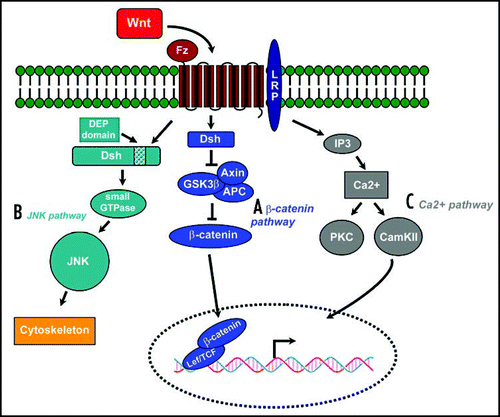
Fig 2 Model of early limb bud initiation in chick (modified from Kawakami et al., 2001). (A) Prior to limb initiation (stage 12), FGF8 (in grey) is expressed in the intermediate mesoderm (IM) adjacent to the presumptive forelimb area. FGF10 (in blue) is expressed in the lateral plate mesoderm (LPM), and in the segmental plate (SP). In the caudal portion of the LPM, FGF10 is coexpressed with Wnt8c (in red). Somites are indicated with their corresponding segment numbers. (B) During limb induction (HH stage 14), both FGF8 (in grey) and Wnt2b (in red) are expressed in the IM. Wnt2b is also expressed in the LPM of the presumptive forelimb area and signals through β-catenin to regulate FGF10 specifically in the prospective forelimb. FGF10 expression remains in the caudal LPM in a diffuse pattern. (C) At HH stage 16, expression of Wnt2b and FGF10 is confined to the forelimb bud field in the LPM. Additionally, expression of FGF10 is now confined to the presumptive hindlimb bud field. It may be Wnt8c in the caudal LPM contributes to this restriction of FGF10. In the LPM of the presumptive limb areas, FGF10 signals to the surface ectoderm (SE) to induce Wnt3a (green) resulting in activation of FGF8. For reason of simplicity, only the expression patterns in the relevant regions are shown, while the expression in somites and other areas is neglected.
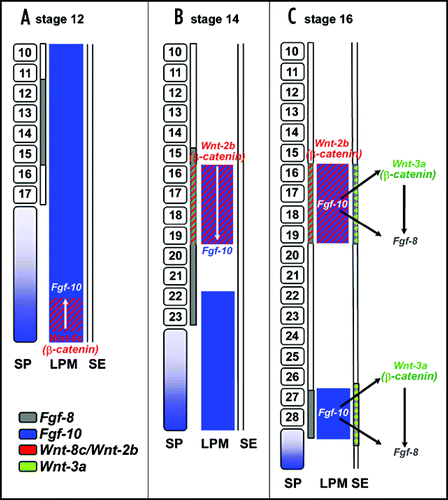
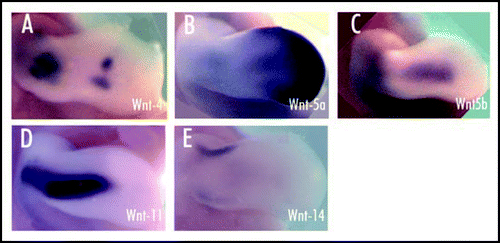
Fig 3 Expression of Wnt genes (4, 5a, 5b, 11, 14) in the mesenchyme surrounding the developing cartilage elements of HH-stage 26 chick forelimb. Limbs shown in dorsal view (Loganathan et al., 2005). (A) Expression of Wnt 4 in the central elbow region and in the joint interzones of the wrist. (B) Wnt 5a expression is seen at the distal tip of the limb bud and in the entire AER, with lower expression levels proximally. (C) Expression of Wnt 5b in the dorsal and ventral mesenchyme of the limb. (D) Wnt 11 transcripts in the dorsal and ventral mesenchyme of the limb. (E) Wnt 14 expression as a transverse stripe in the mesenchyme of the presumptive joint region.