
Abstract
The specification of cell lineages and patterning in the embryo occurs sequentially as specific regions are increasingly restricted in their developmental fates. When and how this occurs is still not entirely clear. Nevertheless, the roles of epigenetic regulatory genes in partitioning the genome into active and inactive domains is evident in a variety of organisms and is highly conserved through evolution. The function of Pax2 in the kidney has been inferred by the phenotypic analysis of loss-of-function mutants in mice, fish, and humans. Although Pax2 and the related gene, Pax8, are essential for early intermediate mesoderm specification and are found in the epithelial lineage arising from that mesoderm, how these proteins regulate cell lineage restriction and gene expression patterns has remained obscure. Our recent data, suggests that Pax proteins help establish chromatin domains within cell lineages by providing the locus and tissue specificity for epigenetic imprinting complexes that modify histones. The novel protein PTIP is a key adaptor that links Pax proteins and possibly many other types of DNA binding proteins to a histone H3K4 methyltransferase complex. Given the prevalence of Pax2 expression in kidney development and in kidney disease, we now need to address the effects of epigenetics on renal disease states, on the stability of the terminal epithelial phenotype, and in the aging cell.
Acknowledgements
I thank Sanj Patel, Ming Fang, Yi Cai for their many contributions in the lab. This work has been supported by NIH grants DK054740 and DK073722 to G.R.D.
Figures and Tables
Figure 1 The mammalian Pax genes. A summary of the nine Pax genes found in mice and humans is outlined. All genes have the characteristic 128 amino acid paired DNA binding domain (PD) at the amino terminus. Based on other conserved sequences, the genes fall into several subfamilies, including the Pax3/7 group which also have a carboxy-terminal homeodomain (HD) and an octapeptide (OD), the Pax4/6 family, which have just the homeodomain, the Pax1/9 family which contains just the octapeptide, and the Pax2/5/8 group which has the octapeptdie and a partial homeodomain. The corresponding mouse and human mutations are indicated as are the key elements of the embryonic expression patterns.
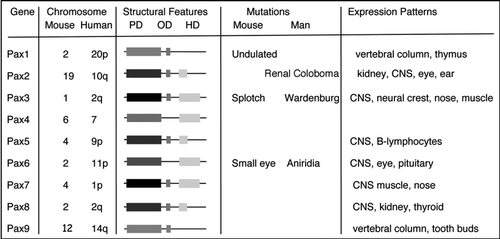
Figure 2 Early patterning of the kidney. (A) In a cross section through a post-gastrulation embryo, the position of the intermediate mesoderm is indicated with respect to the neural tube, notochord, somites and lateral plate. The schematic would correspond to a section posterior to the fourth somite. (B) A longitudinal rendition of the epithelial components from the intermediate mesoderm corresponding to approximately 10.5 days gestation (E10.5) in the mouse. (C) A schematic of the metanephric, or adult, kidney at approximately E12 once the ureteric bud has bifurcated. The cells condensed around the tips of the bud, known as the cap mesenchyme, are the epithelial progenitor cells for the nephrons and become polarized to form primitive renal vesicles.
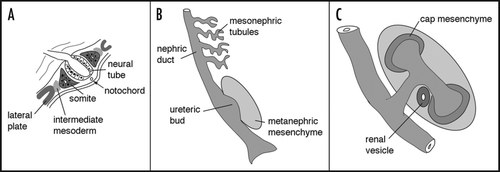
Figure 3 The intermediate mesoderm in Pax2 mutant mice. The region of intermediate mesoderm in wild-type (+/+) and Pax2 homozygous null (−/−) embryos was dissected out and stained with antibodies against cytokeratin, which marks the nephric duct (nd) and Pax2, which marks the metanephric mesenchyme (met) and the mesonephric tubules (mes) tubules. Note the lack of ureteric bud (ub) in Pax2 mutants, even though the nephric duct is initially formed.
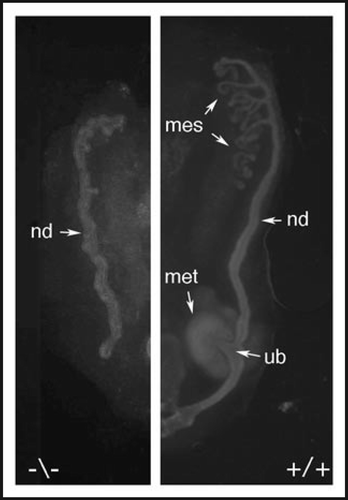
Figure 4 The Pax2 protein is phosphorylated by the c-Jun N-terminal kinase. (A) A schematic of the Pax2 protein showing the critical domains described. The activation domain is required for trans-activation potential in cell culture and contains many proline, serine and threonine residues. (B) The ERK, JNK and p38 kinases were activated in cell culture and used to phosphorylate recombinant GST-Pax2 proteins and control substrates with 32-P. Only the JNK kinase is able to significantly phosphorylate the Pax2 activation domain. (C) The Pax2 activation domain is as good a substrate for recombinant JNK enzyme as the c-Jun protein. (D) Organ cultures derived from E11.5 kidney rudiments and cultured for 48 hours in control media or with 20 µM of the JNK inhibitor SP600125. Cells were stained with antibodies against Pax2, to mark the mesenchyme, and cytokeratins, to mark the branching ureteric buds. Note the lack of condensation of Pax2 positive cells around the ureteric bud tips in the JNK inhibited culture.
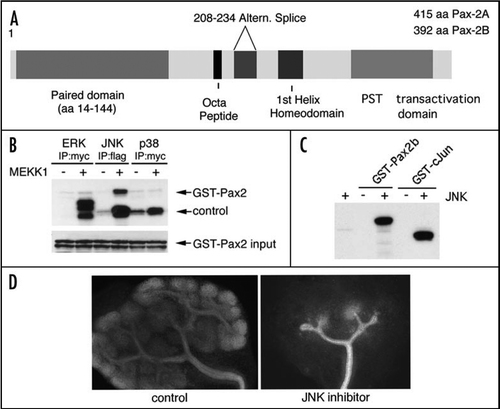
Figure 5 Pax2 and PTIP promote histone methylation. (A) Chromatin immunoprecipitation in cultured cells was done with the indicated antibodies, above the bar graphs and the precipitated DNA analyzed at an integrated Pax2 binding site. Cells were cultured with media alone (1, control), with Pax2 expression plasmids (2, Pax2), with a PTIP siRNA and Pax2 (3, SI Pax2) or rescued with a mouse PTIP that is resistant to the siRNA together with the siRNA and Pax2 (4, SI P + P). Note that Pax2 binds to its recognition sequence regardless of the presence of PTIP. PTIP localizes to the Pax2 recognition site only when Pax2 is expressed but not after knockdown with siPTIP. The H3K4 methyltransferase MLL2 localizes to the Pax2 recognition site in a Pax2 dependent manner and requires PTIP. PTIP knockdowns show no increase in H3K4 dimethylation, but the effect can be rescued with exogenous mouse PTIP. (B) A model of locus specificity for a histone methyltransferase (HMT) complex. Pax2 recognizes a specific sequence and recruits PTIP, potentially through the interaction with Phospho-serines. PTIP provides a link to the HMT complex, resulting in H3K4 methylation. These methylation marks provide docking sites for nucleosome remodeling factors that can promote sliding and assembly of RNA polymerase to activate or maintain gene expression.
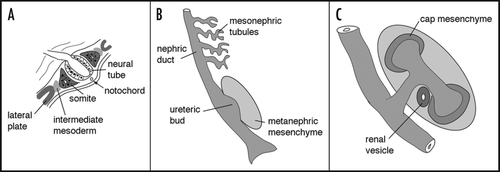
Table 1 Genes that regulate early kidney development and cell lineages
Note
Edited transcripts of research conferences sponsored by Organogenesis and the Washington University George M. O'Brien Center for Kidney Disease Research (P30 DK079333) are published in Organogenesis. These conferences cover organogenesis in all multicellular organisms including research into tissue engineering, artificial organs and organ substitutes and are participated in by faculty at Washington University School of Medicine, St. Louis, MO, USA.