
Abstract
The transcriptional cascade governing adipogenesis has been thoroughly examined throughout the years. Transcription factors PPARγ and C/EBPα are universally recognized as the master regulators of adipocyte differentiation and together they direct the establishment of the gene expression pattern of mature adipose cells. However, this familiar landscape has been considerably broadened in recent years by the identification of novel factors that participate in the regulation of adipogenesis, either favoring or inhibiting it, through their effects on chromatin. Epigenetic signals and chromatin-modifying proteins contribute to adipogenesis and, through regulation of the phenotypic maintenance of the mature adipocytes, to the control of metabolism. In this review we intend to summarize the recently described epigenetic events that participate in adipogenesis and their connections with the main factors that constitute the classical transcriptional cascade.
Acknowledgements
This work was supported by grants BFU2006-14251/BMC and SAF2006-07382 from the Spanish Ministry of Science and Innovation (MICINN) awarded to M.P. and R.G. respectively. We are grateful to Rosa Gasa, Perla Kaliman and Felicia Hanzu for critical reading of the manuscript.
Figures and Tables
Figure 1 Cell fate decision of MSCs. Commitment of MSCs into the chondro-osteogenic lineage has been shown to rely on the expression of the master transcription factor Runx2.Citation114–Citation116 Sox proteins are the major transcription factors that regulate chondrogenic differentiation, whereas Runx2/Cbfa1 and Osterix/Osx participate in both osteocyte and chondrocyte differentiation. The Wnt canonical pathway activates Runx2 and induces an osteogenic fate,Citation117 at the expense of the adipogenic lineage.Citation55,Citation56 On the other hand, BMP2 induces the expression of Sox9 but represses Runx2, thus driving MSCs into chondrogenesis. Commitment to the myogenic lineage and myogenesis are governed by muscle-specific transcription factors that include Myf5, MyoD, myogenin and MRF4,Citation118,Citation119 as well as the induction of TIP-1 expression, whereas expression of TIP-3 directs the progenitors to the adipogenic lineage.Citation59 The myogenic precursors can give rise to either myocytes or brown adipocytes, depending on the absence or presence of adipogenic PRDM16.Citation70,Citation72 However, white adipocytes can also be transdifferentiated into brown adipocytes under different circumstances.
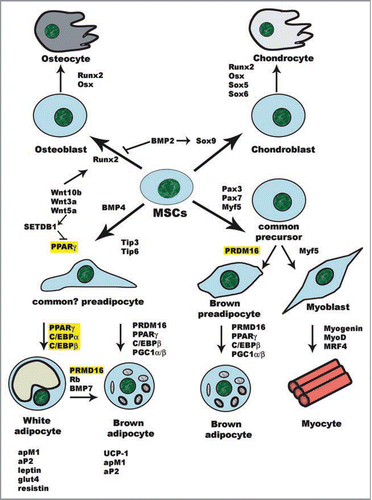
Figure 2 Epigenetic events during adipocyte differentiation. The schematics show the main epigenetic events that take place during adipogenesis represented on a hypothetical promoter incorporating characteristics from different adipogenic genes. In preadipocytes, C/EBPβ is already bound to the promoters of its target genes including pparg and cebpaCitation75,Citation76,Citation80,Citation82 but remains inactive by its interaction with HDAC1/mSin3A.Citation83,Citation84 Similarly, PPARγ is bound to its target promoters in association with a number of repressors including HDACs and histone methyltransferases.Citation57,Citation58,Citation75,Citation76,Citation80–Citation82,Citation97,Citation99 At this stage, most adipogenic genes are characterized by bivalent histone marks, such as methylation of H3K27, H3K9 and H3K4.Citation35–Citation38,Citation50 Methylation of DNA is also foundCitation120 as well as recruitment of a still inactive RNA Pol II.Citation50 Upon induction of differentiation, pRB phosporylation results in its inactivation and HDAC1 is dislodged from PPARγ which is now free to bind CBP/p300 and a number of other coactivators.Citation81,Citation94,Citation95,Citation100 In the early phases of differentiation, increased amounts of PPARγ displace HDAC1 from C/EBPβ allowing the factor to recruit the SWI/SNF remodeling complex and stimulate transcription of cebpa.Citation83 Glucocorticoids have the same effect by stimulating C/EBPβ acetylation by GCN5.Citation84,Citation113 High levels of PPARγ and C/EBPα direct terminal differentiation. Citation75,Citation76 Decrease of H3K27/H3K9 methylation is observed in adipogenic promoters, accompanied by increased histone acetylation and H3K4/H3K20 methylation.Citation38,Citation50,Citation108 Another characteristic feature during terminal differentiation of adipocytes is the spread of di and tri-methylation of H3K4, acetylation of H3 and the Pol II to the coding region of the adipogenic genes, coinciding with high levels of transcription of those genes.Citation38,Citation50
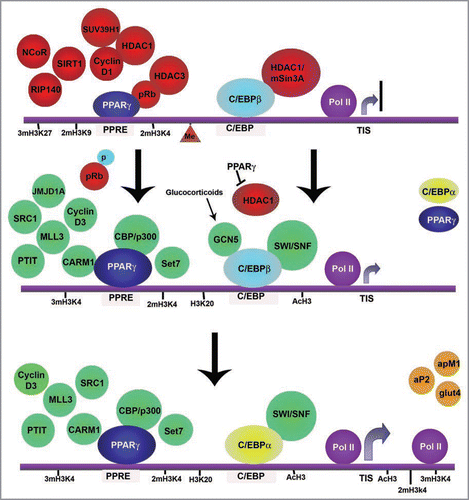
Table 1 Histone modifying enzymes involved in adipogenesis