Abstract
Bidirectional transcription is an interesting feature of eukaryotic genomes; yet not all aspects of its mechanism are understood. Silkmoth choriogenesis is a model system for studying transcriptional regulation at the initiation level. As chorion genes comprise a large group of divergently transcribed gene pairs, we are presented with the possibility of investigating the intricacies of bidirectional transcription. Τheir well characterized 5’ regulatory regions and expression profiles lay the foundation for investigating protein:protein and protein:DNA interactions, and RNA polymerase function during oocyte development. In this article we summarize current knowledge on chorion gene regulation and propose an approach to modeling bidirectional transcription using chorion promoters.
Acknowledgements
We would like to thank Peter R. Cook for comments on the manuscript, and Luc Swevers for discussions on chorion regulation.
Figures and Tables
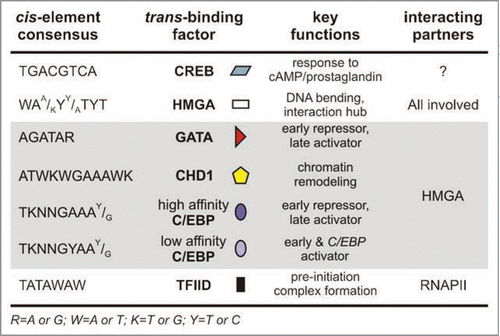
Figure 1 Different kinds of bidirectonal transcription. (A) A pair of divergently transcribed pairs (α- and β-gene; as in chorion gene pairs) sharing a common 5′ regulatory region. Pre-initiation complex (PIC) formation and RNA polymerase II (RNAPII) recruitment precedes transcription, which occurs in both directions (arrows); the number of RNAPII molecules involved and mechanism remain unclear. Two alternative models for this mechanism are present in (C and D) (arrows). (B) A given gene X is under the control of a unidirectional promoter. PIC assembly and RNAPII recruitment close to the transcription start site (TSS) are followed by initiation, mainly producing functional transcripts of X (grey arrow), but occasionally transcribing the antisense strand (white arrows) to produce short non-coding RNA molecules. (C) One or two RNA polymerases per gene pair? Each gene is transcribed separately, in alternate pulses, and RNAPII holo-enzyme complex formation is based on the C/EBP-HMGA-CHD1 complex formed proximal to the α-TATA box. (D) Both genes of the pair are simultaneously transcribed by ‘dedicated’ polymerases, thus formation of two distinct RNAPII complexes—one on each TATA box—is required.
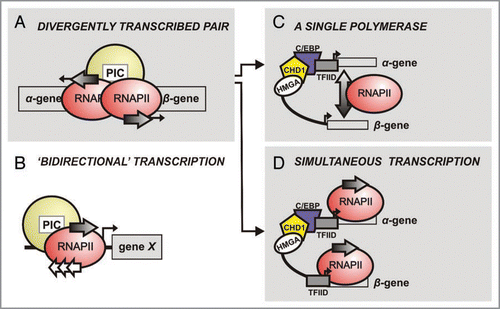
Figure 2 The chorion regulatory circuit. For a chorion gene pair of given developmental specificity, the cascade initiates upon prostaglandin pulses (extra-cellular) triggering second messaging via cAMP. The cAMP response factor CREB is then activated and binds the C/EBP gene promoter to initiate transcription (dotted arrow). C/EBP protein molecules (purple) produced contribute via a positive feedback loop; TFIID recruitment correlates with pre-initiation complex formation. C/EBP is finally switched off as GATA antagonises C/EBP for interaction with the HMGA-CHD1 complex (red line) and binds the promoter. In parallel, C/EBP switches the chorion gene pair on (both α- and β-gene), and proteins produced (grey arrows) integrate to the chorion layer. The target chorion gene is switched off by GATA (red line) and irreversibly ‘muted’ by multiple C/EBP binding events. In all cases, CHD1 acts as the chromatin remodeler to facilitate transcription factor binding.
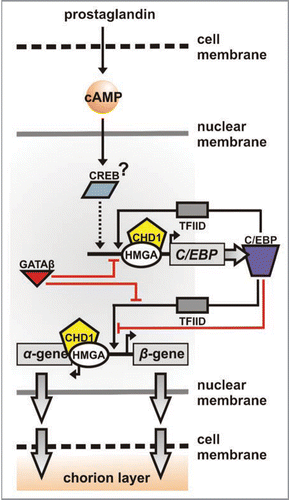
Figure 3 Chorion gene pair promoter regions. Transcription factor binding sites are presented for the early Er.A/B, the early-middle/middle L12-type, the middle/middle-late L11-type and the late Hc.A/B gene promoter regions. Orientation of C/EBP (ovals; light purple, lower affinity; dark purle, higher affinity), and GATA sites (triangles) is denoted. Theoretically predicted CHD1 (pentagons) and HMGA (white boxes) sites are indicated with a questionmark. Distances are drawn to scale (L12-type α- to β-TATA region corresponds to 211 bp); α-proximal arrays of CHD1-HMGA-C/EBP elements masked by a nucleosome appear boxed (red), as do β-proximal ones (green). The gradient bar represents choriogenic periods (E to L) over developmental time (vertical arrow).
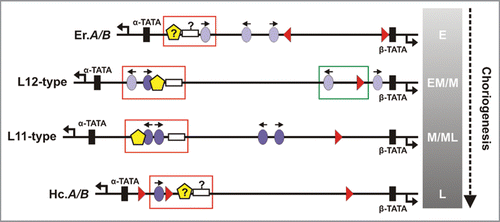
Figure 4 A summary of cis-elements of chorion genes and the protein factors that associate in trans with them. The cis-element consensus sequence, its cognate protein factor, the main function of the factor and proteins it interacts with during choriogenesis are presented. The symbols that accompany each factor are the ones used to depict the relevant binding sites in . Data for the CREB factor are based on binding site prediction and homologies to other insect systems. Symbols used to describe nucleotide composition are explained in the footnote.