Abstract
The increasing prevalence of chronic kidney disease in the absence of new treatment modalities has become a strong driver for innovation in nephrology. An increasing understanding of stem cell biology has kindled the prospects of regenerative options for kidney disease. However, the kidney itself is not a regenerative organ, as all the nephrons are formed during embryonic development. Here, we will investigate advances in the molecular genetics of renal organogenesis, including what this can tell us about lineage relationships, and discuss how this may serve to inform us about both the normal processes of renal repair and options for regenerative therapies.
Acknowledgments
M.L. is a Principal Research Fellow of the National Health and Medical Research Council. I acknowledge that this lecture presented research from past and present members of my research laboratory, including Grant Challen, Michael Lusis, Joan Li, Lorine Wilkinson, Fiona Rae, David Pennisi, Minoru Takasato, Caroline Hendry, Kylie Georgas, Han Sheng Chiu, Jessica Ineson, Norseha Suhaimi, Melissa Becroft and Bree Rumballe, and I thank them for their research efforts. I also acknowledge our collaborators within GUDMAP, notably, Sean Grimmond, Rathi Thiagarajan and Dave Tang at the University of Queensland. Aspects of this work have been supported by the National Institutes of Digestion Diabetes and Kidney, NIH (DK070136, DK070136), the NHMRC (ID45592) and the Australian Stem Cell Centre (P067, Stream 4.2) funding to M.L.
Note
Edited transcripts of research conferences sponsored by Organogenesis and the Washington University George M. O'Brien Center for Kidney Disease Research (P30 DK079333) are published in Organogenesis. These conferences cover organogenesis in all multicellular organisms including research into tissue engineering, artificial organs and organ substitutes and are participated in by faculty at Washington University School of Medicine, St. Louis, MO.
Figures and Tables
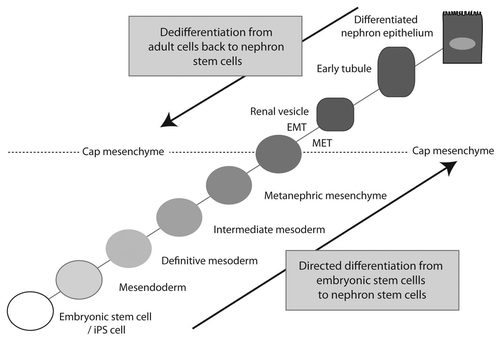
Figure 1 Incomplete lineage map of kidney development. This is based on our current understanding from the mouse. Ovals represent mesenchymal cell types. Rectangles represent epithelial cell types. Solid arrows indicate relationships that are well-supported by the current literature. Dashed lines represent likely relationships without existing lineage tracing evidence. Genes expressed at individual stages are indicated in gray at the appropriate location. Note that the Foxd1+ population gives rise to the mesangial cells, JG cells and vascular pericytes,Citation57 but the direct origin of these cells remains controversial.Citation58 IM, intermediate mesoderm; MM, metanephric mesenchyme; UB, ureteric bud; CM, cap mesenchyme; RV, renal vesicle; CS, connecting segment; DRV, distal RV; PRV, proximal RV; LOH, loop of Henle; JG, juxtaglomerular; GECs, glomerular epithelial cells; PCT, proximal convoluted tubule; PST, proximal straight tubule; DCT, distal convoluted tubule.
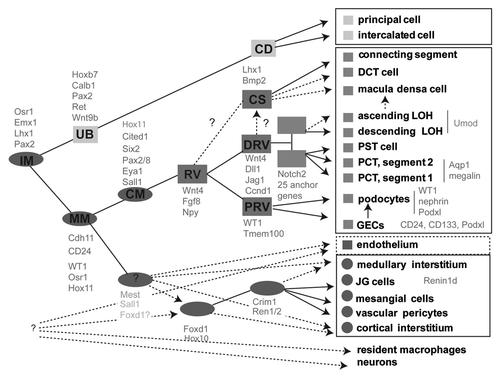
Figure 2 Identification of anchor genes for the early proximal tubule. (A) Heatmap showing genes with the greatest subcompartment specificity. Note the strong subgroup of early proximal tubule (EPT)-specific genes. (B) Intensity plot displaying those genes with the tightest specificity of expression to proximal tubule (PT)/early proximal tubule (EPT) compared with other compartments isolated in Brunskill et al. (C) Validation of specificity of expression of Fbp1 to the proximal tubule from Stage IV of nephron development, expression in developing kidney at E15.5 (top SISH part) and location of the continued expression of this gene in adult kidney (bottom part). (D) Analysis of location of expression of Hdc during development and in adulthood with expression commencing in the earlier Stage III of nephron development (capillary loop stage; top part) and continuing in the adult proximal tubule, notably in segments S1 and S2 (bottom part). Microarray analysis courtesy of Rathi Thiagarajan and Sean Grimmond. SISH data courtesy of Bree Rumballe and Kylie Georgas and available via www.gudmap.org.
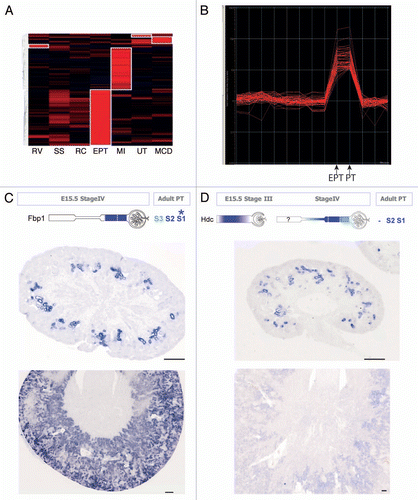
Figure 3 Embryonic kidney nephrospheres and endogenous adult renal mesenchymal stem cells. (A) Nephrospheres cultured from E12.5 embryonic kidney.Citation30 (B) Incorporation of GFP+ nephrosphere-derived cells into the tubule of a developing kidney using recombination explants culture (large arrowhead; tubule outlined with dotted line). (C) Identification of an injected nephrosphere-derived cell in the interstitium of an adult kidney (large arrowhead). (D) Colony-forming unit-fibroblastic (CFU-F) isolated from adult mouse kidney.Citation42 (E) Differential gene expression between bone marrow-derived MS Cs and renal MS Cs. (F–H) Evidence for mesodermal differentiation capacity of renal MS Cs into adipocytes (F), osteocytes (G) and chrondrocytes (H). (I and J) LacZ expression of Crim1 in perivascular cells of the arteries (I), arterioles (I, large arrowhead) and mesangial cells (J, small arrowhead). (K) Expression of Crim1 in the papilla. (l) Model showing the decline in the nephrosphere-forming nephron progenitors toward birth and the increase in prevalence of renal MSCs after birth with respect to other developmental landmarks. Nephrosphere data courtesy of Michael Lusis. MSC data courtesy of Joan Li. Crim1 data courtesy of Lorine Wilkinson.
Figure 4 Section in situ hybridization for the Spp1 (osteopontin) gene in two models of renal injury: unilateral ureteric obstruction (UUO) and ischemia-reperfusion injury (IRI). UUO-CUK, contralateral kidney from animal subjected to UUO; RUUO, reversal of UUO for two weeks following 10 days of obstruction. Left parts are Spp1 SISH images of low magnification sections of adult kidneys. Dotted boxes are numbers to indicate which magnified part they refer to. Columns 2 and 4 show higher magnifications of the cortex (column 2) and papilla (column 4). Column 3 represents dual Spp1 SISH followed by immunohistochemistry for aquaporin 1 (Aqp1) to mark the proximal tubules.Citation59 SISH data courtesy of Bree Rumballe.
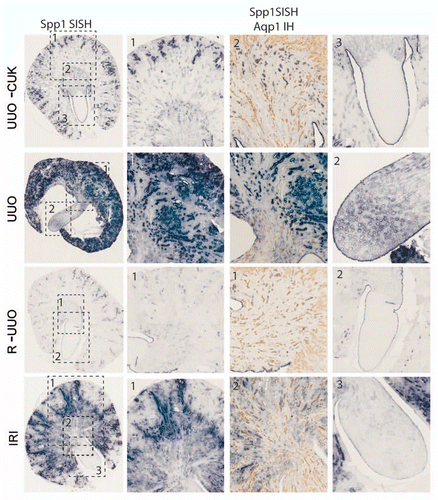
Figure 5 Approaches for the regeneration of nephron progenitor populations. Having molecularly characterized the cap mesenchyme as the source of all cells within the nephron, excepting the collecting duct, this is regarded as the nephron progenitor population. Such a population may be rederived from embryonic stem cells/iPS cells via directed differentiation through the embryological steps originally required to form the cap mesenchyme. Conversely, dedifferentiation from an adult cell type via the enforced re-expression of key transcriptional factor networks may achieve the same endpoint. Cap mesenchyme undergoes an MET to form renal vesicle. If commencing with an epithelial adult cell, one would assume an EMT would be required for the reverse to occur.