Abstract
The serpins are the largest superfamily of protease inhibitors. They are found in almost all branches of life including viruses, prokaryotes and eukaryotes. They inhibit their target protease by a unique mechanism that involves a large conformational transition and the translocation of the enzyme from the upper to the lower pole of the protein. This complex mechanism, and the involvement of serpins in important biological regulatory processes, make them prone to mutation-related diseases. For example the polymerization of mutant α 1-antitrypsin leads to the accumulation of ordered polymers within the endoplasmic reticulum of hepatocytes in association with cirrhosis. An identical process in the neuron specific serpin, neuroserpin, results in the accumulation of polymers in neurons and the dementia FENIB. In both cases there is a clear correlation between the molecular instability, the rate of polymer formation and the severity of disease. A similar process underlies the hepatic retention and plasma deficiency of antithrombin, C1 inhibitor, α 1-antichymotrypsin and heparin co-factor II. The common mechanism of polymerization has allowed us to group these conditions together as a novel class of disease, the serpinopathies.
Acknowledgements
This work was supported by the Medical Research Council (UK), the Wenner Gren Foundations, the Swedish Society for Medical Research and the Isaac Newton Trust Cambridge European Trust.
Figures and Tables
Figure 1 Inhibition of neutrophil elastase by α1-antitrypsin and the structural basis of polymerization. (A) After docking (left) the neutrophil elastase (grey) is inactivated by movement from the upper to the lower pole of the protein (right). This is associated with the insertion of the RCL (red) as an extra strand into β-sheet A (green). (B) The structure of α1-antitrypsin is centred on β-sheet A (green) and the mobile reactive centre loop (red). Polymer formation results from the Z variant of α1-antitrypsin (Glu342Lys at P17; indicated by arrow) or mutations in the shutter domain (blue circle) that open β-sheet A to favour partial loop insertion and the formation of an unstable intermediate (M*). The patent β-sheet A then accepts the loop of another molecule to form a dimer (D), which then extends into polymers (P). The individual molecules of α1-antitrypsin within the polymer, although identical, are coloured red, yellow and blue for clarity. Figure reproduced with permission from Lomas et al.Citation97
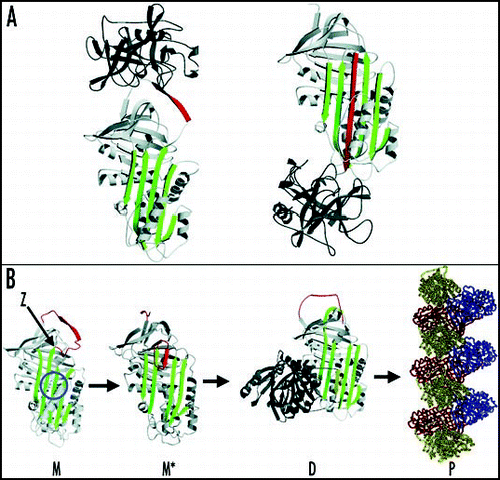
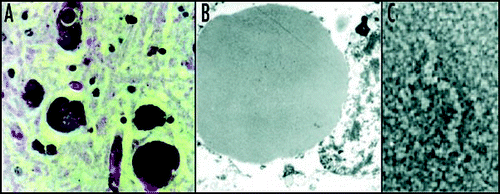
Figure 2 Z α1-antitrypsin is retained within hepatocytes as intracellular inclusions. These inclusions are PAS-positive and diastase resistant (A) and are associated with neonatal hepatitis and hepatocellular carcinoma. (B) Electron microscopy of a hepatocyte from the liver of a patient with Z α1-antitrypsin deficiency shows the accumulation of α1-antitrypsin within the rough ER (arrow). These inclusions are composed of chains of α1-antitrypsin polymers shown here from the plasma of a Siiyama α1-antitrypsin homozygote (C). More recently, polymers have been identified within PAS-positive inclusions with a monoclonal anti-polymer α1-antitrypsin antibody. (D and E) Immunochemistry of the liver from an individual with Z α1-antitrypsin deficiency, showing staining with an anti-α1-antitrypsin polyclonal antibody (D, arrow) and a monoclonal anti-polymer α1-antitrypsin antibody (E, arrow). It is these intracellular inclusions of polymers that are associated with neonatal hepatitis and hepatocellular carcinoma. Figure reproduced with permission from Lomas et al.Citation97
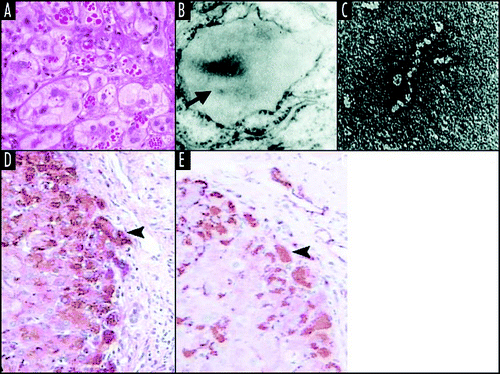
Figure 3 Mutant neuroserpin is retained within neurons as intracellular inclusions. These inclusions stain positive with PAS (A) and can be seen within the ER on electron microscopy (B). Electron microscopy of the isolated inclusions confirms that the mutant neuroserpin forms bead-like polymers identical to those of Z α1-antitrypsin (C). Figure reproduced with permission from Lomas et al.Citation97