
Abstract
The term prion has been used to describe self-replicating protein conformations that can convert other protein molecules of the same primary structure into its prion conformation. Several different proteins have now been found to exist as prions in Saccharomyces cerevisiae. Surprisingly, these heterologous prion proteins have a strong influence on each others’ appearance and propagation, which may result from structural similarity between the prions. Both positive and negative effects of a prion on the de novo appearance of a heterologous prion have been observed in genetic studies. Other examples of reported interactions include mutual or unilateral inhibition and destabilization when two prions are present together in a single cell. In vitro work showing that one purified prion stimulates the conversion of a purified heterologous protein into a prion form, suggests that facilitation of de novo prion formation by heterologous prions in vivo is a result of a direct interaction between the prion proteins (a cross-seeding mechanism) and does not require other cellular components. However, other cellular structures, e.g., the cytoskeleton, may provide a scaffold for these interactions in vivo and chaperones can further facilitate or inhibit this process. Some negative prion-prion interactions may also occur via a direct interaction between the prion proteins. Another explanation is a competition between the prions for cellular factors involved in prion propagation or differential effects of chaperones stimulated by one prion on the heterologous prions.
Acknowledgements
Work in the authors' laboratories was supported by National Science Foundation Grant 0518482 (Irina L. Derkatch) and National Institutes of Health Grant GM056350 (Susan W. Liebman). We thank Yakov Vitrenko, Vidhu Mathur, Andrew O'Dell, Michele Kadnar, Catherine Potenski and N. Kaye Horstman for helpful comments on the manuscript.
Figures and Tables
Figure 1 Cross-seeding model for [PIN+]. [PIN+] aggregates are proposed to be used as the site of initial Sup35 assembly and conversion into [PSI+] (Rnq1, blue arrows; Sup35, green triangles).
![Figure 1 Cross-seeding model for [PIN+]. [PIN+] aggregates are proposed to be used as the site of initial Sup35 assembly and conversion into [PSI+] (Rnq1, blue arrows; Sup35, green triangles).](/cms/asset/ea743660-764f-4911-841a-6584aa95161e/kprn_a_10904837_f0001.gif)
Figure 2 Capping model. The binding of a heterologous protein (magenta diamonds) to the growing tip of a prion aggregate (blue arrows) could block its rapid growth and lead to its destabilization and loss. At the same time such binding can lead to the formation of a prion by the heterologous protein by a seeding mechanism.
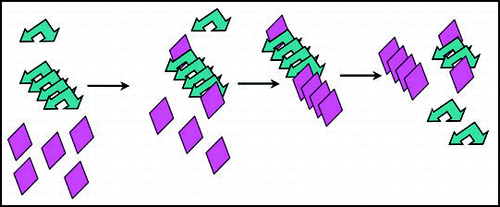
Figure 3 Chaperone-assisted seeding model for [PIN+]. Chaperones (orange sun shapes) bound to preexisting [PIN+] prion aggregates (blue arrows), could be responsible for the enhanced de novo aggregation of Sup35 (green triangles) and thus facilitate [PSI+] formation in vivo.
![Figure 3 Chaperone-assisted seeding model for [PIN+]. Chaperones (orange sun shapes) bound to preexisting [PIN+] prion aggregates (blue arrows), could be responsible for the enhanced de novo aggregation of Sup35 (green triangles) and thus facilitate [PSI+] formation in vivo.](/cms/asset/6ff1b909-100c-4e53-a710-69b2188e3eee/kprn_a_10904837_f0003.gif)
Figure 4 Titration model for [PIN+]. Top: in a [psi-] [pin-] cell, cellular factors (purple shape) keep Sup35 (green triangles) and Rnq1 (blue arrows) from aggregating. Bottom: in a [psi-] [PIN+] cells, much of the factor is bound to the [PIN+] aggregate, so less is available to keep Sup35 from aggregating.
![Figure 4 Titration model for [PIN+]. Top: in a [psi-] [pin-] cell, cellular factors (purple shape) keep Sup35 (green triangles) and Rnq1 (blue arrows) from aggregating. Bottom: in a [psi-] [PIN+] cells, much of the factor is bound to the [PIN+] aggregate, so less is available to keep Sup35 from aggregating.](/cms/asset/52629559-c68f-4031-8620-822aca35b83d/kprn_a_10904837_f0004.gif)
Figure 5 Cellular response model to explain why [PSI+] destabilizes [URE3]. Top: normal propagation of [URE3] (red pentagons). Bottom: [PSI+] (linked green triangles) induces the expression of the Ssa1 chaperone (light blue sickles), which destabilizes [URE3] but not [PSI+].
![Figure 5 Cellular response model to explain why [PSI+] destabilizes [URE3]. Top: normal propagation of [URE3] (red pentagons). Bottom: [PSI+] (linked green triangles) induces the expression of the Ssa1 chaperone (light blue sickles), which destabilizes [URE3] but not [PSI+].](/cms/asset/a1245f12-3858-4dab-9491-305486076c57/kprn_a_10904837_f0005.gif)
Notes
This manuscript has been previously published: Derkatch IL, Liebman, SW. Prion-Prion Interactions. In: Protein-Based Inheritence. Chernoff, Y ed. Austin and New York: Landes Bioscience and Kluwer Academic Press, 2007; 39–46.