Abstract
Amyloid formation is a hallmark of several systemic and neurodegenerative diseases. Extracellular amyloid deposits or intracellular inclusions arise from the conformational transition of normally soluble proteins into highly ordered fibrillar aggregates. Amyloid fibrils are formed by nucleated polymerization, a process also shared by prions, proteinaceous infectious agents identified in mammals and fungi. Unlike so called non-infectious amyloids, the aggregation phenotype of prion proteins can be efficiently transmitted between cells and organisms. Recent discoveries in vivo now implicate that even disease-associated intracellular protein aggregates consisting of α-synuclein or Tau have the capacity to seed aggregation of homotypic native proteins and might propagate their amyloid states in a prion-like manner. Studies in tissue culture demonstrate that aggregation of diverse intracellular amyloidogenic proteins can be induced by exogenous fibrillar seeds. Still, a prerequisite for prion-like propagation is the fragmentation of proteinaceous aggregates into smaller seeds that can be transmitted to daughter cells. So far efficient propagation of the aggregation phenotype in the absence of exogenous seeds was only observed for a yeast prion domain expressed in tissue culture. Intrinsic properties of amyloidogenic protein aggregates and a suitable host environment likely determine if a protein polymer can propagate in a prion-like manner in the mammalian cytosol.
Acknowledgements
We are thankful to M. Suhre and T. Scheibel (Universität Bayreuth, Germany) for providing a Sup35 plasmid and the production of rec. Sup35NM and S. Kremmer (Helmholtz Zentrum München, Germany) for generating NM antibodies. We thank the members of the Vorberg and Schätzl laboratories for constructive comments on this work. Financial support for this work was provided by the Deutsche Forschungsgemeinschaft (SFB 596 B14, VO 1277/1-2) and by the EU NoE Neuroprion.
Figures and Tables
Figure 1 Schematic representation of amyloid and prion formation. (A) Assembly of amyloid fibrils follows a nucleated polymerization reaction with a lag phase required to form an aggregation nucleus followed by an elongation phase in which monomers are rapidly recruited into growing aggregates. Oligomers are common folding intermediates of amyloid fibrils. Amyloid fibrils have a cross-β structure, bind to fluorophore thioflavine T and exhibit Congo Red birefringence. (B) Efficient prion propagation follows the nucleated polymerization process. Spontaneous induction of prion protein nuclei is a rare event. In yeast, the prion phenotype [PIN+] of the host protein Rnq1 strongly enhances the de novo appearance of other prions (e.g., the prion conformation of Sup35, [PSI+]), likely by cross-seeding. Induction mechanisms in spontaneous human prion diseases are unknown. The disaggregase Hsp104 is essential for partitioning large prion polymers into smaller propagons that are transmitted during cell division or mating in yeast. Essential co-factors for mammalian prion propagation remain elusive.
![Figure 1 Schematic representation of amyloid and prion formation. (A) Assembly of amyloid fibrils follows a nucleated polymerization reaction with a lag phase required to form an aggregation nucleus followed by an elongation phase in which monomers are rapidly recruited into growing aggregates. Oligomers are common folding intermediates of amyloid fibrils. Amyloid fibrils have a cross-β structure, bind to fluorophore thioflavine T and exhibit Congo Red birefringence. (B) Efficient prion propagation follows the nucleated polymerization process. Spontaneous induction of prion protein nuclei is a rare event. In yeast, the prion phenotype [PIN+] of the host protein Rnq1 strongly enhances the de novo appearance of other prions (e.g., the prion conformation of Sup35, [PSI+]), likely by cross-seeding. Induction mechanisms in spontaneous human prion diseases are unknown. The disaggregase Hsp104 is essential for partitioning large prion polymers into smaller propagons that are transmitted during cell division or mating in yeast. Essential co-factors for mammalian prion propagation remain elusive.](/cms/asset/e06c72c0-4630-4b25-9a72-d1aba17452f0/kprn_a_10910013_f0001.gif)
Figure 2 Propagation of NM-HA aggregates in mammalian cells. (A) N2a cells stably expressing NM-HA were treated with in vitro generated fibrils of recombinant Sup35NM and subsequent passages were analyzed by confocal microscopy. Once induced, NM-HA aggregates were efficiently transmitted to daughter cells during cell division. Arrows mark cells that undergo cell division. (B) NM-HA is soluble when stably expressed in HpL3-4 cells. (C) Induction and propagation of NM-HA aggregates in HpL3-4 cells. Two passages after treatment with recombinant Sup35NM fibrils, HpL3-4_NM-HA cells exhibit NM-HA aggregates (marked by arrows). Antibody, anti-HA. Nuclei were visualized using Hoechst dye.
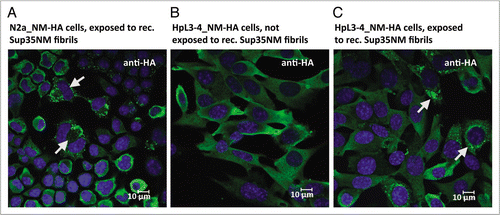
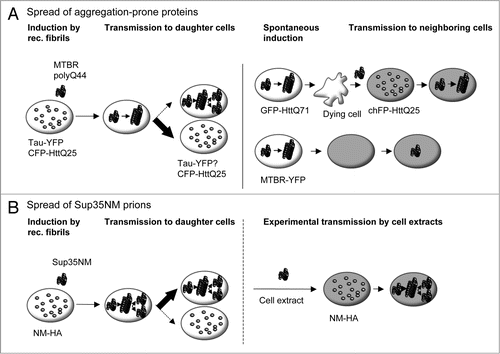
Figure 3 Comparison of aggregation and dissemination propensities of cytosolic aggregation-prone proteins and Sup35NM prions in cell culture. (A) Induction of homotypic protein aggregates and transfer of aggregates to recipient cells. Left, yellow-fluorescent protein-tagged wild-type Tau (Tau-YFP) and cyan-fluorescent protein tagged Huntingtin fragment HttQ25 (CFP-HttQ25) exhibit low spontaneous aggregation propensities.Citation44,Citation45 Uptake of fibrillar aggregates of recombinant polyglutamine peptides (polyQ44) or the microtubule binding region of Tau (MTBR) triggers co-aggregation with soluble cognate proteins. Induced CFP-HttQ25 aggregates display low mitotic stability. Maintenance of wild-type Tau aggregates has not been assessed. Right: GFP-HttQ71 aggregates formed spontaneously upon transient expression and released by dying cells can be taken up by co-cultured recipient cells where they trigger aggregation of soluble cherry fluorescent protein-tagged HttQ25 (chFP-HttQ25). Recipient cells are marked in grey. Similarly, YFP-tagged MTBR aggregates (MTBR-YFP) spontaneously formed upon transient expression in cell culture are transferred to recipient cells. (B) Sup35NM prion propagation. Spontaneous formation of NM-HA aggregates is a very rare event.Citation52 Aggregation of NM-HA can be initiated by recombinant Sup35NM fibrils.Citation43 Once induced, the prion phenotype is maintained during cell division probably due to a very effective mechanism of aggregate fractionation and dissemination. Cell-derived Sup35NM prions are experimentally transmissible to mammalian cells expressing soluble NM-HA and to a lower extent to yeast. Mechanisms of cell-to-cell transmission in cell culture are currently under investigation.