
Abstract
In eukaryotic cells amyloid aggregates may incorporate various functionally unrelated proteins. In mammalian diseases this may cause amyloid toxicity, while in yeast this could contribute to prion phenotypes. Insolubility of amyloids in the presence of strong ionic detergents, such as SDS or sarcosyl, allows discrimination between amorphous and amyloid aggregates. Here, we used this property of amyloids to study the interdependence of their formation in yeast. We observed that SDS-resistant polymers of proteins with extended polyglutamine domains caused the appearance of SDS or sarcosyl-insoluble polymers of three tested chromosomally-encoded Q/N-rich proteins, Sup35, Rnq1 and Pub1. These polymers were non-heritable, since they could not propagate in the absence of polyglutamine polymers. Sup35 prion polymers caused the appearance of non-heritable sarcosyl-resistant polymers of Pub1. Since eukaryotic genomes encode hundreds of proteins with long Q/N-rich regions, polymer interdependence suggests that conversion of a single protein into polymer form may significantly affect cell physiology by causing partial transfer of other Q/N-rich proteins into a non-functional polymer state.
Acknowledgements
The work was supported by the grants from Russian Foundation for Basic Research (# 08-04-0062) and the Wellcome Trust. Authors are grateful to M. Sherman for providing the plasmid p103Q-GFP and to O. Mitkevich for the help in obtaining antibody against Pub1. V.N.U. and A.B.V. contributed equally to this work.
Figures and Tables
Figure 1 Schematic representation of the proteins used in this work. N, M, C, Sup35 domains. 2HA, double hemagglutinin tag. F, FLAG tag. Grey bars, amyloidogenic regions; Q/N, Q/N-rich; Q, Q-rich. Black bars, RNA-binding domains of Pub1.
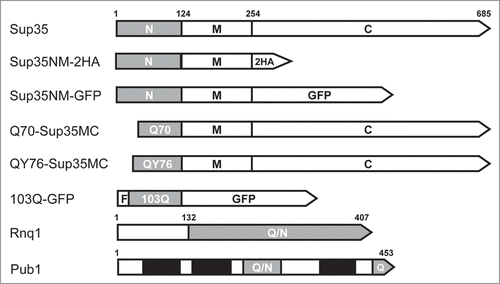
Figure 2 Polymers of Q70 and QY76 cause polymerization of the plasmid-encoded Sup35NM-2HAprotein in Δrnq1 cells. (A) The plasmid pRS315-SUP35C of the strain 74-D694ΔS35 [psi−][PIN+] was shuffled for either pQ70-Sup35MC or pQY76-Sup35MC. Then, the RNQ1 gene was deleted. Polymers of Q70 and QY76 were revealed using SDD-AGE and western blotting with polyclonal anti-Sup35NM antibody. (B) These strains were transformed with a multicopy plasmid, encoding Sup35NM-2HA. Appearance of Sup35NM-2HApolymers was tested using SDD-AGE and western blotting with monoclonal anti-HA antibody. PolyQ, the Q70 or QY76 protein; −70, −76, transformants that have lost the plasmid encoding indicated polyQ protein. Sup35NM-2HA, − or +, absence or presence of the plasmid encoding this protein. RNQ1, +, chromosomal wild type allele, Δ, disruption allele.
![Figure 2 Polymers of Q70 and QY76 cause polymerization of the plasmid-encoded Sup35NM-2HAprotein in Δrnq1 cells. (A) The plasmid pRS315-SUP35C of the strain 74-D694ΔS35 [psi−][PIN+] was shuffled for either pQ70-Sup35MC or pQY76-Sup35MC. Then, the RNQ1 gene was deleted. Polymers of Q70 and QY76 were revealed using SDD-AGE and western blotting with polyclonal anti-Sup35NM antibody. (B) These strains were transformed with a multicopy plasmid, encoding Sup35NM-2HA. Appearance of Sup35NM-2HApolymers was tested using SDD-AGE and western blotting with monoclonal anti-HA antibody. PolyQ, the Q70 or QY76 protein; −70, −76, transformants that have lost the plasmid encoding indicated polyQ protein. Sup35NM-2HA, − or +, absence or presence of the plasmid encoding this protein. RNQ1, +, chromosomal wild type allele, Δ, disruption allele.](/cms/asset/884b9d91-4694-439c-b64d-3d45de32c52e/kprn_a_10911074_f0002.gif)
Figure 3 Polymers of Q70 and QY76 cause aggregation of Sup35NM-GFP in cells lacking [PIN+]. Multicopy plasmids encoding Q70 or QY76 were introduced into 74-D694 [psi−][PIN+] cells and then RNQ1 was deleted to remove [PIN+]. After this, the plasmid encoding Sup35NM-GFP was introduced. About 25% of transformed cells contained fluorescent dots. The same strain, but without the pQ70-Sup35MCand pQY76-Sup35MCplasmids was used as a control.
![Figure 3 Polymers of Q70 and QY76 cause aggregation of Sup35NM-GFP in cells lacking [PIN+]. Multicopy plasmids encoding Q70 or QY76 were introduced into 74-D694 [psi−][PIN+] cells and then RNQ1 was deleted to remove [PIN+]. After this, the plasmid encoding Sup35NM-GFP was introduced. About 25% of transformed cells contained fluorescent dots. The same strain, but without the pQ70-Sup35MCand pQY76-Sup35MCplasmids was used as a control.](/cms/asset/7fd6d28c-4751-45e9-9194-794ef7fcfc2c/kprn_a_10911074_f0003.gif)
Figure 4 Time-dependent appearance of the 103Q-GFP and Sup35 polymers upon induction of 103Q-GFP synthesis. Cells of the strain 74-D694 [psi−][PIN+] with the multicopy p103Q-GFP plasmid were grown in glucose-containing medium (Gl), then in raffinose-containing medium (R), and then transferred to galactose-containing medium (Gal) and incubated for 1, 1.5, 2 and 3 h. After this, the 103Q-GFP-encoding plasmid was lost (−). (A) Polymers of 103Q-GFP and (B) Sup35 revealed by SDD-AGE. (C) The levels of polymer, monomer and total Sup35, SDS-PAGE analysis. The samples in the upper panel were not boiled before loading onto the gel. In the middle of the run, the gel was taken out, boiled to dissolve Sup35 polymers and allowed to run further. Blots were stained with anti-GFP (A) and anti-Sup35NM (B and C) antibodies.
![Figure 4 Time-dependent appearance of the 103Q-GFP and Sup35 polymers upon induction of 103Q-GFP synthesis. Cells of the strain 74-D694 [psi−][PIN+] with the multicopy p103Q-GFP plasmid were grown in glucose-containing medium (Gl), then in raffinose-containing medium (R), and then transferred to galactose-containing medium (Gal) and incubated for 1, 1.5, 2 and 3 h. After this, the 103Q-GFP-encoding plasmid was lost (−). (A) Polymers of 103Q-GFP and (B) Sup35 revealed by SDD-AGE. (C) The levels of polymer, monomer and total Sup35, SDS-PAGE analysis. The samples in the upper panel were not boiled before loading onto the gel. In the middle of the run, the gel was taken out, boiled to dissolve Sup35 polymers and allowed to run further. Blots were stained with anti-GFP (A) and anti-Sup35NM (B and C) antibodies.](/cms/asset/8bec769a-8092-4a72-bf95-a9fa375bbeaa/kprn_a_10911074_f0004.gif)
Figure 5 Co-appearance of SDS-resistant polymers of Q70, Rnq1 and Pub1. Cells of the 74-D694ΔS35 [psi−][pin−-] strain were transformed with the pQ70-SUP35MC plasmid. Samples were taken at 30 (lane 1) and 100 (lane 2) cell generations after transformation. Then the pQ70-SUP35MC plasmid was replaced with pRS315-SUP35C (lane 3). The samples were analyzed by SDD-AGE and western blotting. The proteins indicated below were revealed using antibodies to Sup35NM, Rnq1 and Pub1, respectively.
![Figure 5 Co-appearance of SDS-resistant polymers of Q70, Rnq1 and Pub1. Cells of the 74-D694ΔS35 [psi−][pin−-] strain were transformed with the pQ70-SUP35MC plasmid. Samples were taken at 30 (lane 1) and 100 (lane 2) cell generations after transformation. Then the pQ70-SUP35MC plasmid was replaced with pRS315-SUP35C (lane 3). The samples were analyzed by SDD-AGE and western blotting. The proteins indicated below were revealed using antibodies to Sup35NM, Rnq1 and Pub1, respectively.](/cms/asset/286369e7-aecc-4b94-858c-2a8d350e67d6/kprn_a_10911074_f0005.gif)
Figure 6 Pub1 polymers depend on the [PSI+] determinant. (A) Polymers of Sup35 and Pub1 in [PSI+][PIN+] cells, (B) Absence of Pub1 polymers in [psi−][PIN+] cells, (C) Disappearance of Pub1 polymers upon [PSI+] elimination, (D) Pub1 can polymerize in [psi−][PIN+] cells overproducing Rnq1. + and −, presence or absence of a prion determinant; −*, [PSI+] was eliminated by Hsp104 overproduction; wt and ↑, wild type or increased levels of Pub1; +↑ and +M, Rnq1 was overproduced in [PIN+] cells from multicopy plasmids with either the native or MET17 promoter. Note: Pub1 monomers occupy a large area at the bottom of a gel. This is likely to be due to a high monomer proportion for Pub1 and low resolution of agarose gel for monomeric proteins aggravated by overloading of this area of the gel, which accommodates most of the cellular proteins.
![Figure 6 Pub1 polymers depend on the [PSI+] determinant. (A) Polymers of Sup35 and Pub1 in [PSI+][PIN+] cells, (B) Absence of Pub1 polymers in [psi−][PIN+] cells, (C) Disappearance of Pub1 polymers upon [PSI+] elimination, (D) Pub1 can polymerize in [psi−][PIN+] cells overproducing Rnq1. + and −, presence or absence of a prion determinant; −*, [PSI+] was eliminated by Hsp104 overproduction; wt and ↑, wild type or increased levels of Pub1; +↑ and +M, Rnq1 was overproduced in [PIN+] cells from multicopy plasmids with either the native or MET17 promoter. Note: Pub1 monomers occupy a large area at the bottom of a gel. This is likely to be due to a high monomer proportion for Pub1 and low resolution of agarose gel for monomeric proteins aggravated by overloading of this area of the gel, which accommodates most of the cellular proteins.](/cms/asset/55e114b5-79d2-4800-b1d3-7ee3299ddef6/kprn_a_10911074_f0006.gif)
Table 1 Plasmids