Abstract
Prion diseases are fatal neurodegenerative disorders caused by prion proteins (PrP). Infectious prions accumulate in brain through a template mediated conformational conversion of endogenous PrPC into alternately folded PrPSc. Immunoassays toward pre-clinical detection of infectious PrPSc have been confounded by low-level prion accumulation in non-neuronal tissue and the lack of PrPSc selective antibodies. We report a method to purify infectious PrPSc from biological tissues for use as an immunogen and sample enrichment for increased immunoassay sensitivity. Significant prion enrichment is accomplished by sucrose gradient centrifugation of infected tissue and isolation with detergent resistant membranes from lipid rafts (DRMs). At equivalent protein concentration a 50-fold increase in detectable PrPSc was observed in DRM fractions relative to crude brain by direct ELISA. Sequential purification steps results in increased specific infectivity (DRM >20-fold and purified DRM immunogen >40-fold) relative to 1% crude brain homogenate. Purification of PrPSc from DRM was accomplished using phosphotungstic acid protein precipitation after proteinase-K (PK) digestion followed by size exclusion chromatography to separate PK and residual protein fragments from larger prion aggregates. Immunization with purified PrPSc antigen was performed using wild-type (wt) and Prnp0/0 mice, both on Balb/cJ background. A robust immune response against PrPSc was observed in all inoculated Prnp0/0 mice resulting in antisera containing high-titer antibodies against prion protein. Antisera from these mice recognized both PrPC and PrPSc, while binding to other brain-derived protein was not observed. In contrast, the PrPSc inoculum was non-immunogenic in wt mice and antisera showed no reactivity with PrP or any other protein.
Acknowledgements
This work was supported and administered under the USDA-ARS National Program in Animal Health (CRIS #5325-32000-008-00D), by a USDA Cooperative Agreement (58-5325-3-246), by grants from the National Institutes of Health (AG02132, AG10770 and AG021601) and by a gift from the G. Harold and Leila Y. Mathers Charitable Foundation.
Figures and Tables
Figure 1 Isolation of prions from crude brain homogenate with detergent resistant membranes (DRM). Sucrose density gradient centrifugation separates lipid-associated proteins from the majority of brain proteins via differences in sucrose buoyancy at 4°C. Fractionation of the gradient (12 × 1 mL) resulted in detection of a small protein peak observed within the 5–30% sucrose zone (buoyant fractions #4–5), whereas the bulk of proteins from both PrPC and PrPSc brain homogenate were retained at the bottom of the gradient (A). Western blots revealed that the majority of detectable PrPC and PrPSc were localized to the lipid-rich DRM fractions #4–5 (B; top parts). Proteinase-K (+PK) digestion of the gradient fractions from PrPSc infected brain resulted in an expected molecular weight shift and detection of PrPSc in the DRM fractions (B; top right part). The DRM protein marker Flotillin-1 served as a positive control to confirm the integrity of the gradient and localization of the DRM fractions (B; bottom parts). The majority of PrPC and proteinase-K resistant PrPSc were detected by ELISA in DRM fractions #4–5 (C).
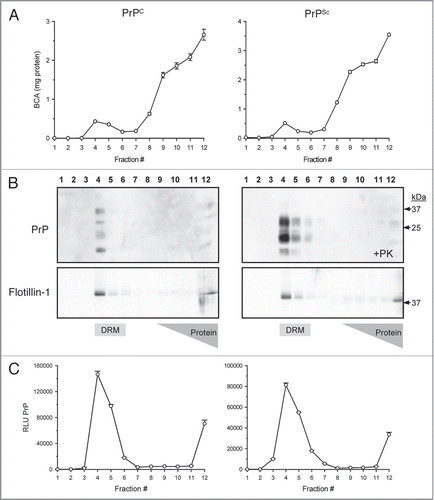
Figure 2 Enrichment of prions in detergent resistant membrane fractions (DRM). Increased detection of PrP 27–30 in DRM fraction (>40-fold) relative to crude brain homogenate by direct ELISA (A). Protein normalization by BCA (5 µg/mL); RLU = relative light units; quantitation of three independent samples (Mean ± SE M; p =<0.001). Western blot comparison of detectable PrP 27–30 in crude hamster brain homogenates, DRM and phosphotungstic acid (PTA) precipitated DRM proteins (B). Progressive increase in detection of three PrPSc glycoforms in DRM fractions and PTA precipitated DRM fractions compared to crude brain homogenates. Protein normalization by BCA prior to PK-treatment and PTA precipitation.
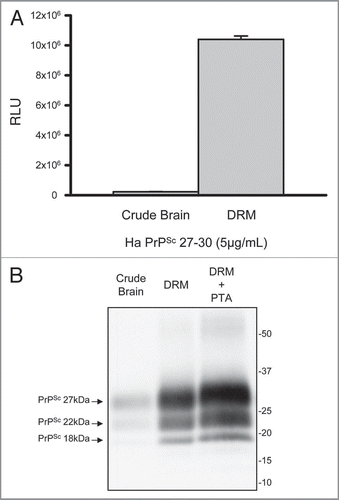
Figure 3 Prion resides as a high molecular weight aggregate in detergent resistant membrane (DRM) fractions. Western blot detection of PrPC following fractionation of normal brain DRM on a linear sucrose gradient showed detection of PrPC across a range of molecular weights composed primarily of mono- and di-glycosylated isoforms (top left part). The oligomeric status of PrPSc from Scrapie infected DRM differs from PrPC with the majority detected as high molecular weight (Fraction #12; >440 kDa) aggregate (top right part). This high molecular weight PrPSc species is composed primarily of mono- and di-glycosylated PrP with detection of non-glycosylated PrP across lower molecular weights (Fractions #3–10). Proteinase-K digestion of gradient fractions from Scrapie DRM shows that all the PK-resistant PrPSc is localized to fraction #12 as a high molecular weight aggregate whereas the non-glycosylated PrP in fractions #3-10 is PK-sensitive (bottom right part). Caveolin-1 (Cav-1) and flotillin-1 (Flot-1) are established DRM protein markers known to form high molecular weight oligomeric complexes and were used as positive controls to validate the integrity of the sucrose gradient (middle and bottom left parts respectively). The linear sucrose gradient was calibrated with know molecular weight protein standards and their mobility and molecular weight are indicated by the large arrows at the bottom of .
Figure 4 Concentration and purification of prion by PTA and size exclusion chromatography. Enriched PrPSc DRM fractions were treated with proteinase-K then proteins PTA precipitated, the solubalized (PrPSc DRM-PK-PTA) material was then fractionated by size exclusion chromatography (Sephadex G100). Protein concentration of fractions was determined by BCA assay (A; solid line) and PrPSc detection by ELISA (A; dashed line). A small protein peak was observe in fraction #5 that corresponded to the void fraction of the column (proteins >100 kDa) which also contained the majority of detectable PrPSc. Western blot detection of G100 fractionated PrPSc DRM-PK-PTA-G100 showed a major band in the void fraction #5 (B). A second PTA protein precipitation following G100 fractionation recovered detectable PrPSc in the void fractions (C). Evaluation of the purified PrPSc DRMPK-PTA-G100-PTA material by silver and Coomassie stain (D; left and middle part respectively) showed detectable PrPSc protein at the expected molecular weight that corresponded to PrPSc detection by Western blot (D; right part).
Figure 5 Increasing specific infectivity with PrPSc purification. Days of survival were determined by hamster bioassay for purified Scrapie brain homogenate preparations (A). Transmissible disease was observed following intracerebral inoculation of 1% PrPSc (25 µg), PrPSc DRM (1.35 µg) and PrPSc PK-treated PTA precipitated material fractionated by size exclusion on Sephadex G100 (PrPSc DRM-PK-PTA-G100; 0.8 µg). N = sample size. Comparison of ID50 and specific infectivity following intracerebral inoculation of 1% crude brain homogenate (brain), DRM and purified prion (DRM-PK-PTA-G100) by incubation time assay (B). Isolation of PrPSc in DRMs from lipid rafts results in >20-fold and purified PrPSc DRM-PK-PTA-G100 >40-fold, increase in specific infectivity relative to crude brain.
Figure 6 Schematic diagram of prion purification and immunization strategy. An outline of steps involved in prion purification from brain homogenate to immunization of Prnp0/0/Balbc/J and wild-type Balbc/J mice with purified PrPSc (PrPSc DRM-PK-PTA-G100-PTA).
Figure 7 Detection of SH aPrP (90-231) by ELISA with antisera from prion immunized Prnp0/0/Balbc/J, but not wt Balbc/J mice. Antisera from Prnp0/0/Balbc/J mice detected SH aPrP (90-321) at dilutions >1:30 K by direct binding ELISA; whereas antisera from immunized wt Balbc/J mice failed to detect SH aPrP (90-231) at any dilution (A). Antisera from prion immunized Prnp0/0/Balbc/J, but not wt mice, detected SH aPrP (90-231), Syrian hamster brain PrPC (Ha PrPSc DRM; no PK) and PrPSc (Ha PrPSc DRM; +PK) by Western blot (B). Detection of PK-resistant PrPSc was observed with antisera diluted >25 K from two representative Prnp0/0/Balbc/J (#1–2) and wt Balbc/J (#3–4) mice immunized with purified prion antigen. RLU = relative light units.
Figure 8 Antisera from Prnp0/0/Balbc/J mice immunized with purified PrPSc binds Syrian hamster prion proteins. Western blot comparing binding of antisera from a prion imunized Prnp0/0/Balbc/J to normal and infectious hamster brain homogenate (30 µg/lane), recSH aPrP(90-231); 100 ng/lane) and normal and infectious hamster brain DRM (10 µg/lane) preparations (top part). Antisera detected PK-sensitive PrPC, recSH aPrP(90-231) and PK-insensitive PrPSc in brain homogenates and DRM preparations; no other protein bands were observed. A comparative western blot showed similar binding of the monoclonal anti-prion antibody IPC1 to brain homogenate, recombinant PrP and DRM preparations (bottom part) as antisera from Prnp0/0/Balbc/J mice. PK = proteinase-K treatment (+). Protein normalization by BCA.
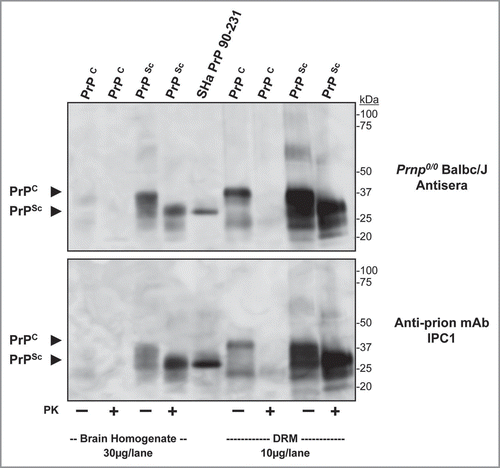
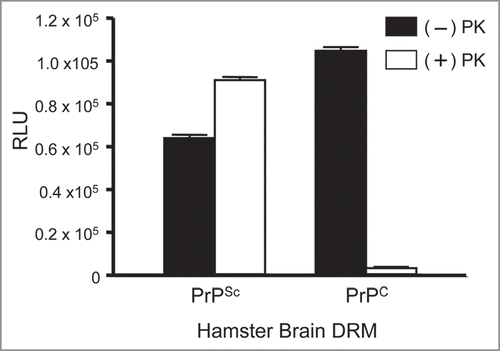
Figure 9 Detection of endogenous prion proteins in normal (PrPC) and diseased (PrPSc) hamster brain DRM fractions by ELISA. Antisera from a prion immunized Prnp0/0/Balbc/J mouse (1:10K) bound DRM fraction from both PrPC and PrPSc in the absence of PK (−); black bars. Detection of PK-resistant PrP27-30 was observed in PrPSc, but not PrPC, DRM fractions following PK-treatment (+); open bars. No binding to any DRM fraction was observed with antisera from non-immunized Prnp0/0/Balbc/J mice. ELISA performed in triplicate and DRM protein normalized to 2 µg/well by BCA. RLU = relative light units. PK = proteinase-K.