
Abstract
Despite intensive research into how amyloid structures can impair cellular viability, the molecular nature of these toxic species and the cellular mechanisms involved are not clearly defined and may differ from one disease to another. We systematically analyzed, in Saccharomyces cerevisiae, genes that increase the toxicity of an amyloid (M8), previously selected in yeast on the sole basis of its cellular toxicity (and consequently qualified as “artificial”). This genomic screening identified the Vps-C HOPS (homotypic vacuole fusion and protein sorting) complex as a key-player in amyloid toxicity. This finding led us to analyze further the phenotype induced by M8 expression. M8-expressing cells displayed an identical phenotype to vps mutants in terms of endocytosis, vacuolar morphology and salt sensitivity. The direct and specific interaction between M8 and lipids reinforces the role of membrane formation in toxicity due to M8. Together these findings suggest a model in which amyloid toxicity results from membrane fission.
Acknowledgements
We gratefully acknowledge Johanna Coindreau for her technical help and enthusiasm in searching for KO strains showing changes in M8 and WT sensitivity. This work was supported by grants from the CNRS (Programme Interdisciplinaire—Interface Physique Chimie Biologie—Soutien à la prise de Risque) and from the French Research National Agency ANR grant, project ANR-06-MRAR-011-01 «AMYLOI». K.B. received fellowships from the ANR and the Conseil Régional d'Aquitaine.
Figures and Tables
Figure 1 Viability assays for WT and KO strains. Growth of BY4742 or mutant cells expressing GFP (▴), WT (◆) or M8 (▪). Cells were grown overnight on dextrose medium supplemented with casaminoacids (0.67%). When cells reached the exponential phase, they were transferred into galactose medium supplemented with casaminoacids (0.05 OD600 nm/ml). Cell concentrations were evaluated by OD600 nm at various times after the induction over a period of 120 h.
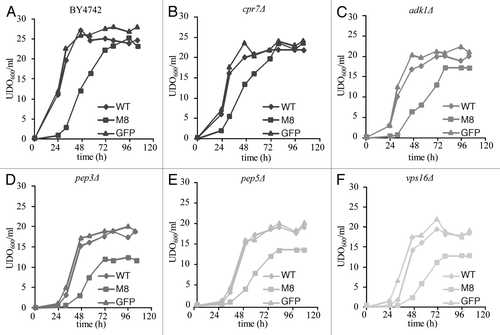
Figure 2 Metal and salt sensitivity spotting assay. Ten-fold dilutions of exponentially growing cultures of BY4742 cells expressing GFP, WT or M8 were spotted onto SD agar supplemented with casaminoacids (0.67%) and containing 8 mM MnCl2, 100 mM MgCl2, 25 mM LiCl, 100 mM CaCl2 or 10 mM citric acid (A). pep3Δ, pep5Δ and vps16Δ expressing GFP or WT were tested in the same conditions (B). The cells were incubated at 30°C for 4 days.
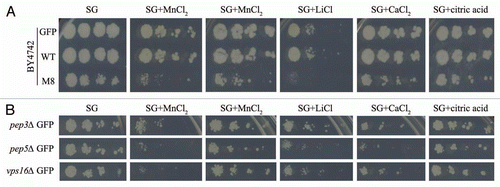
Figure 3 Vacuolar morphology observation by microscopy. Exponentially growing cultures of BY4742 cells expressing GFP, WT or M8 were incubated for 15 min on ice with the fluorescent marker FM 4–64 (20 µM). Then, the cells were washed twice and were incubated for 45 min at 30°C. Cells were mounted in medium and observed with a DMRB microscope with a 100X HCX PL fluotar objective.
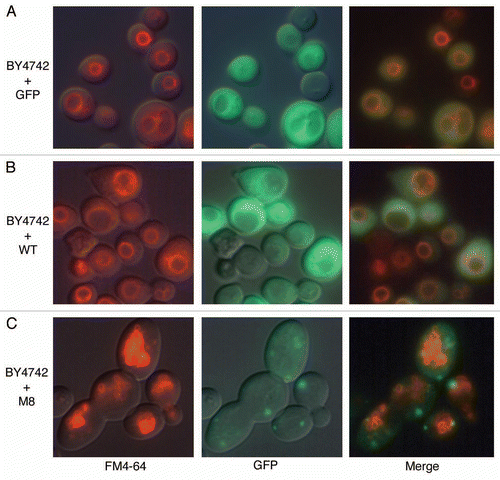
Figure 4 M8 interacts specifically with lipids in vitro. M8 interacts with DMPC blotted onto a PVDF membrane. M8 is detected directly by Ponceau red and indirectly by an anti-histidine antibody.
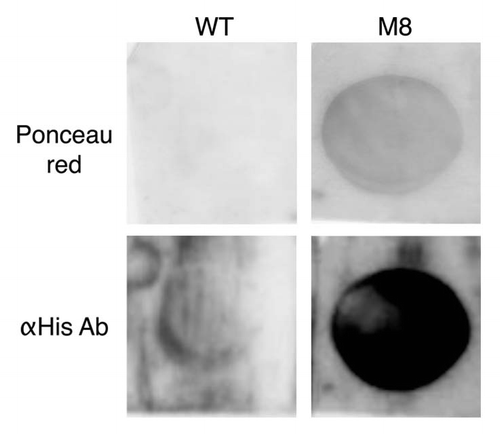
Figure 5 Traffic monitoring. (A) Ten-fold dilutions of exponentially growing cultures of BY4742 cells expressing GFP, WT or M8 were spotted onto SG agar with or without 5 µM 5-FU. Petri dishes were observed after 10 days at 30°C to allow the observation of M8 growth. (B) Exponentially growing cultures of BY4742 or mutant cells expressing GFP or M8 were used to prepare total cell extracts. The samples were run on a 12% SDS-PAGE gel, transferred onto a nitrocellulose membrane and exposed to anti-CPY antibodies. m, mature protein; p, precursor protein.
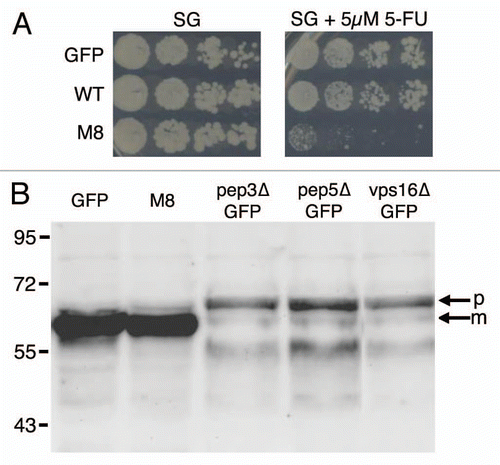
Figure 6 M8 affects vesicle trafficking. The class C HOPS complex promotes small vesicles, like early endosomes, fusion to the vacuole. M8 may prevent this fusion by coating the vacuole with an hydrophobic layer of proteins. Both M8 expression and the absence of the class C HOPS complex will affects vesicle formation leading to an accumulation of small vesicles and to a significantly greater growth impairment. Target vesicles may be vacuolar vesicles or may be other endocytosis vesicles.
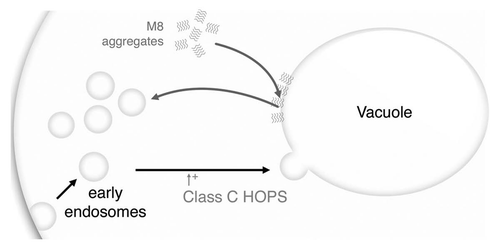
Table 1 MIPS and GO classification of verified M8 toxicity enhancers