
Abstract
The infectious agent of prion diseases is believed to be nucleic acid-free particles composed of misfolded conformational isomers of a host protein known as prion protein (PrP). Although this “protein-only” concept is generally accepted, decades of extensive research have not been able to elucidate the mechanisms by which PrP misfolding leads to neurodegeneration and infectivity. The challenges in studying prion diseases relate in part to the limitations of mammalian prion models, which include the long incubation period post-infection until symptoms develop, the high expense of maintaining mammals for extended periods, as well as safety issues. In order to develop prion models incorporating a genetically tractable simple system with a well-defined neuronal system, we generated transgenic C. elegans expressing the mouse PrP behind the pan-neuronal ric-19 promoter (Pric-19). We show here that high expression of Pric-19::PrP in C. elegans can result in altered morphology, defective mobility, and shortened lifespan. Low expression of Pric-19::PrP, however, does not cause any detectable harm. Using the dopamine neuron specific promoter Pdat-1, we also show that expression of the murine BAX, a pro-apoptotic member of the Bcl-2 family, causes dopamine neuron destruction in the nematode. However, co-expression of PrP inhibits BAX-mediated dopamine neuron degeneration, demonstrating for the first time that PrP has anti-BAX activity in living animals. Thus, these distinct PrP-transgenic C. elegans lines recapitulate a number of functional and neuropathological features of mammalian prion models, and provide an opportunity for facile identification of genetic and environmental contributors to prion-associated pathology.
Acknowledgements
The authors thank I. Ruvensky for the gift of pRIC19 plasmid; D. Harris for the gifts of pESC-LeuGAL10-mBax and p426GPD-mBI-HA; J. Richmond and Anna Burdina for technical assistance; J. Kramer, J. Richmond and R. Nass for helpful discussions; members of the Li laboratory for critical comments; and E. Crow, R. Nass and C. Long for manuscript editing. This work was supported by grants from the United States Army (0850-370-R744), the Ellison Medical Foundation and the US National Institutes of Health (R01NS056086) to L.L.
Figures and Tables
Figure 1 Creating transgenic C. elegans lines that express mouse prion protein (PrP). (A) Shown are Pric-19::GFP and Pric-19::PrP constructs, which contain the green fluorescence protein (green box) and the full-length PrP (1–254) (grey box) including the indicated N-terminal signal sequence (1–22) and the C-terminal site (231) for GPI anchoring, respectively. The ric-19 promoter (blue arrow) is also indicated. (B) After co-injection of the Pric-19::GFP and Pric-19::PrP constructs into the gonads of healthy young adult worms, five integrant lines expressing both GFP and PrP were obtained (see Materials and Methods). Shown are immunoblot analyses of the five lines using a PrP specific monoclonal antibody 3F4 and anti-actin. An integrant line expressing GFP alone was included as control.
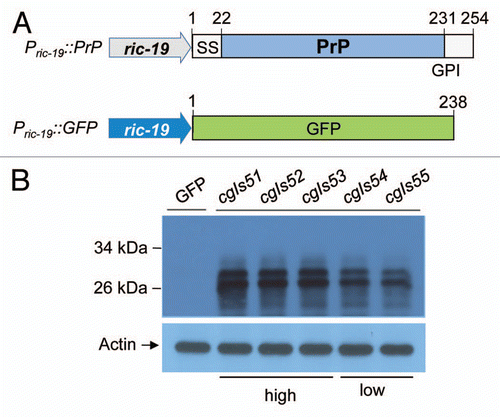
Figure 2 PrP molecules produced in C. elegans were glycosylated, GPI-anchored and presented on the plasma membranes of neurons. (A) Crude cell lysates prepared from high (cgIs51) and low (cgIs55) expressers were treated with (+) or without (−) PNGase F, which removes N-linked oligosaccharide chains from glycoproteins, followed by immunoblot analysis using the antibody 3F4. (B) Crude lysates from cgIs51 and cgIs55 were also treated with (+) or without (−) phosphatidylinositol-specific phospholipase C (PIPLC) for 2 h at 4°C, which releases GPI-anchored PrP molecules from the detergent phase, D, to the aqueous phase, A. After Triton X-114 treatment and phase partitioning, proteins were precipitated and immunoblotted with 3F4 antibody. (C) Nematodes co-expressing Pric-19::PrP and Pric-19::GFP were freeze-crack treated, incubated with the 3F4 antibody then a TRITC-labeled secondary antibody and followed by photographing under a fluorescence microscope. Left, PrP immunostaining; middle, ric-19::GFP expression; right, a merged image showing that GFP was localized in the cytoplasm whereas PrP was on the cell surface surrounding the GFP signal. Upper: high expresser line cgIs51; Lower: low expresser line cgIs55. The length of the bars in each image is 25 µm. The inset shows an enlarged motor neuron cell body on the ventral nerve cord (arrow heads).
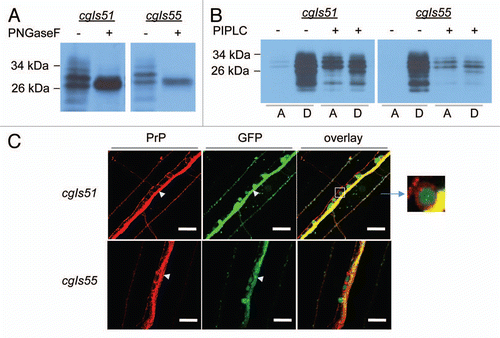
Figure 3 Morphologic and behavioral characteristics of transgenic worms expressing PrP in neurons. (A) Visualization of GFP expression in neurons of high (cgIs51 and cgIs53) and low (cgIs55) expressing lines of Pric-19::PrP/Pric-19::GFP. A transgenic worm expressing only Pric-19::GFP (GFP) was included as a control. The length of the bar in each image is 50 µm. As shown, the high expressers had dumpy-like body morphology: short and chunky-looking. (B) The high expressers (cgIs51 and cgIs53) also exhibited decreased lifespan whereas the low expressers (cgIs55) showed a lifespan comparable to that of non-transgenic control (N2) and worms expressing GFP only. Average lifespan in days (±SD) are: N2, 22.6 ± 2.5; GFP, 21.1 ± 1.8; cgIs51, 12.3 ± 1.8; cgIs53, 10.82 ± 2.5 and cgIs55, 19.3 ± 2.7. (C) Locomotion measured by liquid thrashing assays at adult stage for N2, GFP, cgIs51, cgIs53 and cgIs55worms. Each data point is the mean thrashing rate for ten worms. (D) Pharyngeal pumping rates were analyzed by counting terminal bulb contractions for a 20 sec period. (E) pBOC cycles were assayed on food by measuring the time between two posterior body contractions. For each assay, ten animals were used for each genotype and their average values are shown. *p < 0.01, in all cases, compared with the non-transgenic worms (N2) by Student's t-test.
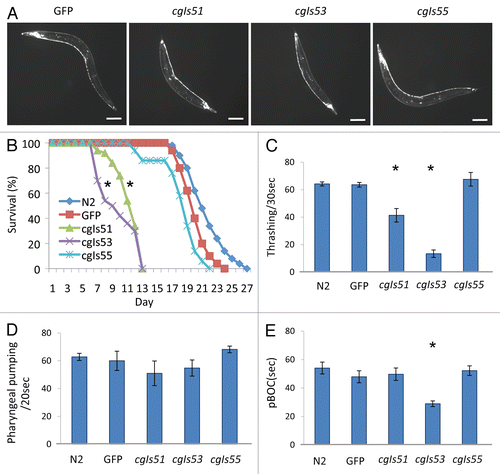
Figure 4 BAX expression in C. elegans dopamine neurons results in a comprised food sensibility and PrP inhibits BAX-mediated food sensibility reduction. (A) Food sensing behavioral assays of transgenic worms with their genotypes indicated, including two integrant lines of Pdat-1::BAX. The locomotion rates of 3-day-old worms were counted in the presence (with food) or absence (no food) of bacteria lawn. (B) Food sensing assay performed in worms coexpressing both PrP in whole neurons (Pric-19 based) and BAX in dopamine neurons (Pdat-1 based). (C) Food sensing assay in worms coexpressing PrP and BAX in dopamine neurons. *p < 0.01 by Student's t-test.
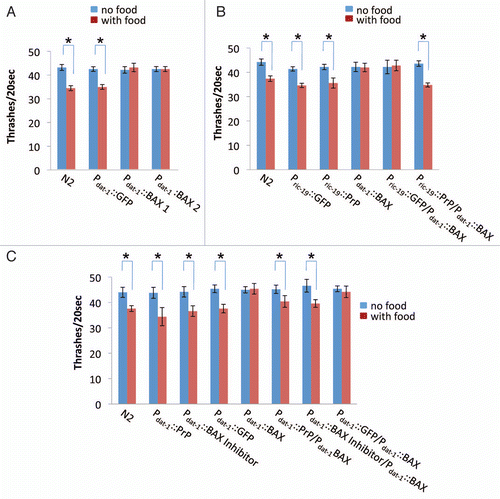
Figure 5 PrP inhibits BAX-induced cell destruction in C. elegans dopamine neurons. (A) Fluorescence microscopic examination of the dopamine neurons of living worms expressing Pdat-1::GFP. Worms with or without BAX, co-expressing PrP or BI-1 were examined and their CEP and ADE neurons are shown (arrows). All transgenes were expressed under Pdat-1. The length of the bar in each image is 25 µm. (B) Immunoblot analyses showing that PrP, BAX inhibitor and BAX were expressed in the integrant lines of cgIs56[dat-1(BAX)], cgIs58[dat-1(PrP)] and cgIs59[dat-1(BI)], respectively. (C) Food sensing assay carried out in worms co-expressing BAX and cytosolic PrP or BAX and anchorless PrP in dopamine neurons. *p < 0.01 by Student's t-test.
![Figure 5 PrP inhibits BAX-induced cell destruction in C. elegans dopamine neurons. (A) Fluorescence microscopic examination of the dopamine neurons of living worms expressing Pdat-1::GFP. Worms with or without BAX, co-expressing PrP or BI-1 were examined and their CEP and ADE neurons are shown (arrows). All transgenes were expressed under Pdat-1. The length of the bar in each image is 25 µm. (B) Immunoblot analyses showing that PrP, BAX inhibitor and BAX were expressed in the integrant lines of cgIs56[dat-1(BAX)], cgIs58[dat-1(PrP)] and cgIs59[dat-1(BI)], respectively. (C) Food sensing assay carried out in worms co-expressing BAX and cytosolic PrP or BAX and anchorless PrP in dopamine neurons. *p < 0.01 by Student's t-test.](/cms/asset/551693a7-c28a-496c-bfd6-c8a17c549826/kprn_a_10914026_f0005.gif)
Table 1 Integrant C. elegans strains used for this study