Abstract
The prion diseases occur following the conversion of the cellular prion protein (PrPC) into a disease-related isoform (PrPSc). In this study a cell painting technique was used to examine the role of the glycosylphosphatidylinositol (GPI) anchor attached to PrPC in prion formation. The introduction of PrPC to infected neuronal cells increased the cholesterol content of cell membranes, increased activation of cytoplasmic phospholipase A2 (cPLA2) and increased PrPSc formation. In contrast, PrPC with a monoacylated GPI anchor did not alter the amount of cholesterol in cell membranes, was not found within lipid rafts and did not activate cPLA2. Although monoacylated PrPC remains within cells for longer than native PrPC it was not converted to PrPSc. Moreover, the presence of monoacylated PrPC displaced cPLA2 from PrPSc-containing lipid rafts, reducing the activation of cPLA2 and PrPSc formation. We conclude that acylation of the GPI anchor attached to PrPC modifies the local membrane microenvironments that control some cell signaling pathways, the trafficking of PrPC and PrPSc formation. In addition, such observations raise the possibility that the pharmacological modification of GPI anchors might constitute a novel therapeutic approach to prion diseases.
Acknowledgments
This work was supported by a grant from the European Commission FP6 “Neuroprion” Network of Excellence.
Figures and Tables
Figure 1 Phospholipase digestion of PrPC affects the acylation of the GPI anchor. Cartoon showing the putative GPI anchor attached to PrPC, monoacylated PrPC and deacylated PrPC. Glycan residues shown include inositol (Inos), mannose (Man), sialic acid (SA), galactose (Gal), N-acetyl galactosamine (GalNAc) and glucosamine (GlcN) as well as phosphate (P).
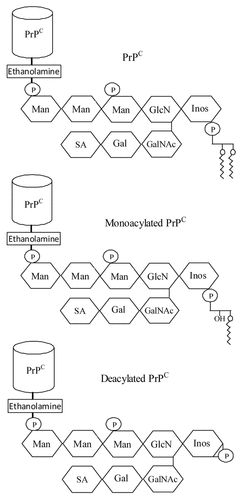
Figure 2 Acylation of PrPC affects the underlying cell membrane. Cartoon showing the proposed membranes surrounding native PrPC and monoacylated PrPC, including cholesterol (), lyso-phospholipids (![]()
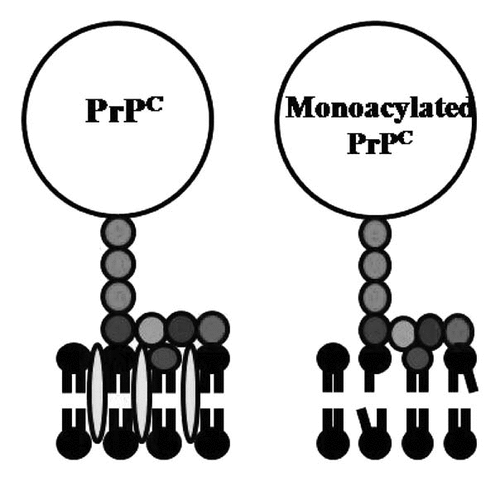
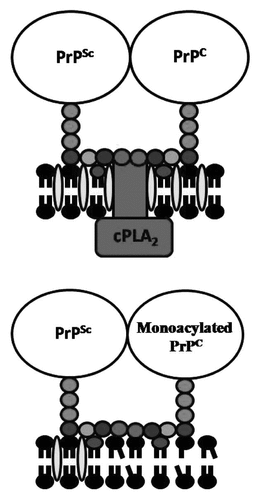
Figure 3 Monoacylated PrPC affects the capture of cPLA2 in PrPSc-containing lipid rafts. (A) Cartoon showing the proposed membranes surrounding PrPSc and PrPC including the capture of cPLA2 in lipid rafts that are dense in cholesterol (![]()