
Abstract
Protein misfolding and assembly into ordered, self-templating aggregates (amyloid) has emerged as a novel mechanism for regulating protein function. For a subclass of amyloidogenic proteins known as prions, this process induces transmissible changes in normal cellular physiology, ranging from neurodegenerative disease in animals and humans to new traits in fungi. The severity and stability of these altered phenotypic states can be attenuated by the conformation or amino-acid sequence of the prion, but in most of these cases, the protein retains the ability to form amyloid in vitro. Thus, our ability to link amyloid formation in vitro with its biological consequences in vivo remains a challenge. In two recent studies, we have begun to address this disconnect by assessing the effects of the cellular environment on traits associated with the misfolding of the yeast prion Sup35. Remarkably, the effects of quality control pathways and of limitations on protein transfer in vivo amplify the effects of even slight differences in the efficiency of Sup35 misfolding, leading to dramatic changes in the associated phenotype. Together, our studies suggest that the interplay between protein misfolding pathways and their cellular context is a crucial contributor to prion biology.
Acknowledgments
We thank A. Derdowski for the FLIP image and members of the Serio lab for helpful discussions and comments on the manuscript. This work was supported by awards from the National Institutes of Health to T.R.S. (GM069802) and to S.D. (AG032818).
Note Added in Proof
Recent studies by the Weissman group have determined that amyloid fibers composed of G58D alone, when present in a particular conformation, exhibit the same conversion and thermodynamic stability differences in comparison with wild-type fibers in vitro as we have identified for mixed G58D-wild-type prion aggregates in vivo.Citation140
Figures and Tables
Figure 1 The cellular environment modulates prion misfolding pathways to create protein-based traits. (top) Self-replicating protein conformations create stable phenotypes (white colonies) in vivo when the processes of synthesis (1), conversion (2), fragmentation (3) by Hsp104 (hexamer) and transmission (4) are balanced to allow aggregates of prion proteins to persist in vivo. (middle) Overexpression of a prion protein promotes the conversion reaction (red arrows), leading to the accumulation of large aggregates that are inefficiently transmitted to daughter cells (dotted red arrow) and loss of the prion-associated phenotype (red colonies). (bottom) Dominant inhibition of prion propagation by mutants that decrease conversion efficiency (dotted red arrow) or enhance fragmentation efficiency (solid red arrow) promote aggregate disassembly (ball and stick) and induce prion loss (red colonies).
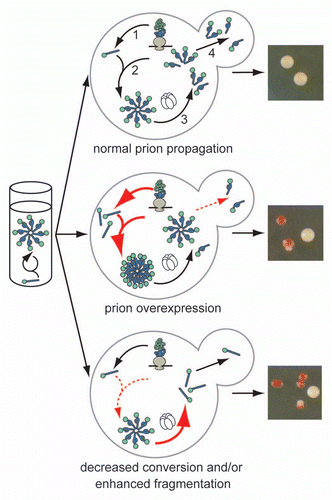
Figure 2 Relating prion phenotypic severity to aggregate thermodynamic stability. For many prion variants, there is a linear but inverse relationship between aggregate thermodynamic stability and phenotypic severity (black dotted line), but this trend cannot explain the phenotypes associated with all prion variants or the effects of dominant-negative prion mutants (see text for details). Our studies in vivo on the yeast prion [PSI+] suggest that thermodynamic stability poses a limit on prion persistence at both extremes (red line) by impacting aggregate size and accumulation (shown schematically). The least thermodynamically stable aggregates are efficiently resolubilized, while the most thermodynamically stable aggregates are inefficiently transmitted.
![Figure 2 Relating prion phenotypic severity to aggregate thermodynamic stability. For many prion variants, there is a linear but inverse relationship between aggregate thermodynamic stability and phenotypic severity (black dotted line), but this trend cannot explain the phenotypes associated with all prion variants or the effects of dominant-negative prion mutants (see text for details). Our studies in vivo on the yeast prion [PSI+] suggest that thermodynamic stability poses a limit on prion persistence at both extremes (red line) by impacting aggregate size and accumulation (shown schematically). The least thermodynamically stable aggregates are efficiently resolubilized, while the most thermodynamically stable aggregates are inefficiently transmitted.](/cms/asset/56823a06-0c44-444e-9b76-72fec2b5b72a/kprn_a_10916413_f0002.gif)
Figure 3 Transmission of Sup35 protein to daughter cells is conformation-dependent. (left) Schematic of fluorescence loss in photobleaching assay (FLIP) for Sup35 transmission to daughter cells. Bleached daughter (red) and monitored mother (black) are indicated. (right) Fluorescence retention in mother cells expressing Sup35-GFP in the [PSI+]strong (white), [PSI+]weak (pink) or [psi−] (red) conformation.
![Figure 3 Transmission of Sup35 protein to daughter cells is conformation-dependent. (left) Schematic of fluorescence loss in photobleaching assay (FLIP) for Sup35 transmission to daughter cells. Bleached daughter (red) and monitored mother (black) are indicated. (right) Fluorescence retention in mother cells expressing Sup35-GFP in the [PSI+]strong (white), [PSI+]weak (pink) or [psi−] (red) conformation.](/cms/asset/5ef91a15-3bc9-44c0-b8c9-92c0bc548768/kprn_a_10916413_f0003.gif)