
Abstract
Yeast prions are heritable protein-based genetic elements which rely on molecular chaperone proteins for stable transmission to cell progeny. Within the past few years, five new prions have been validated and 18 additional putative prions identified in Saccharomyces cerevisiae. The exploration of the physical and biological properties of these “nouveau prions” has begun to reveal the extent of prion diversity in yeast. We recently reported that one such prion, [SWI+], differs from the best studied, archetypal prion [PSI+] in several significant ways.1 Notably, [SWI+] is highly sensitive to alterations in Hsp70 system chaperone activity and is lost upon growth at elevated temperatures. In that report we briefly noted a correlation amongst prions regarding amino acid composition, seed number and sensitivity to the activity of the Hsp70 chaperone system. Here we extend that analysis and put forth the idea that [SWI+] may be representative of a class of asparagine-rich yeast prions which also includes [URE3], [MOT3+] and [ISP+], distinct from the glutamine-rich prions such as [PSI+] and [RNQ+]. While much work remains, it is apparent that our understanding of the extent of the diversity of prion characteristics is in its infancy.
Financial Support
This work was supported by the National Institute of Health Grant GM31107 (E.A.C.) and National Institute of Health NRSA postdoctoral fellowship GM846542 (J.K.H.).
Note added in proof
A recent publication by the Lindquist laboratory (Halfmann, et al. Opposing effects of glutamine and asparagine govern prion formation by intrinsically disordered proteins. Mol Cell 2011; 43:72–84) has provided additional support for the idea presented herein that amino acid composition of a PrD, particularly the ratio of N to Q residues, affects both mitotic stability and the propensity for de novo prion formation.
Figures and Tables
Figure 1 The Cyclic Hsp70 Chaperone System. Ssa (purple), the yeast cytosolic Hsp70, binds and releases client polypeptides (blue) in a regulated and ATP-dependent manner. J-proteins (aquamarine) including Sis1, Ydj1 and others, stimulate Ssa ATP hydrolysis by virtue of a conserved J-domain and thereby catalyze the “forward” direction of the cycle as indicated above. ADP•Ssa more stably associates with client polypeptides than the ATP-bound form and hence J-proteins favor the ADP•Ssa•Peptide complex. In some cases, J-proteins can also bind and deliver client polypeptides to Hsp70s via C-terminal domains (also shown above). Nucleotide exchange factors (NEFs), including the Sse proteins (dark blue) which share some structural homology with Ssa, catalyze the “reverse” direction of the cycle by facilitating ADP release and subsequent ATP binding, and thus favor an ATP•Ssa state with a dissociated peptide.
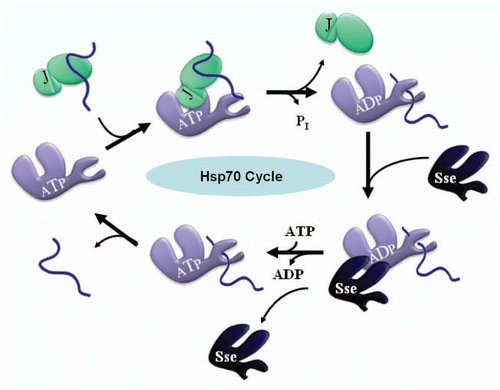
Table 1 Sensitivity of prions to ectopic chaperone expression
Table 2 Correlations amongst yeast prions