Abstract
Stress signaling is central to plants which—as immobile organisms—have to endure environmental fluctuations that constantly interfere with vigorous growth. As a result, plant-specific, elaborate mechanisms have evolved to perceive and respond to stress conditions. Currently, these stress responses are plausibly being revealed to involve crosstalks with energy signaling pathways as any growth-limiting factor alters plant’s energy status. Among these, autophagy, conventionally regarded as the mechanism whereby plants recycle and remobilize nutrients in bulk, has frequently been associated with stress responses. With the recent discoveries, however, autophagy has attained a novel role in stress signaling. In this review, major elements of abitoic stress signaling are summarized along with autophagy pathway, and in the light of recent discoveries, a putative, state-of-art role of autophagy is discussed.
Keywords: :
Text
Due to their sessile nature, plants are constantly subjected to multiple environmental stresses to varying extents. Consequently, plants are equipped with elegant response mechanisms involving several stress-inducible gene products, to enable them to maintain homeostasis against these stresses.Citation1-Citation4 Stress-inducible genes are generally classified into two major groups: functional proteins that provide direct tolerance to abiotic stresses (such as LEA proteins, enzymes that are involved in osmoprotectant biosynthesis) and regulatory proteins (mainly transcription factors, such as DREBs, AREBs and NACs) responsible for downstream signal transduction and induction of functional proteins.Citation3,Citation5 The phytohormone abscisic acid (ABA) has been a major focus of research into stress response mechanisms in plants. In addition to its pivotal roles in vegetative growth, seed dormancy and stomatal aperture, ABA is involved as a central mediator in several stress responses and thus has been considered as a “stress hormone.”Citation1,Citation4-Citation8 Plant water status is tightly related to ABA levels; water deficit resulting from drought or salinity stresses triggers ABA accumulation, which in turn triggers response mechanisms via several ABA-responsive genes, ultimately leading to physiological changes such as the rapid closure of stomata and synthesis of osmoprotectants.Citation1,Citation8
Plants respond to drought and salinity stresses via both ABA-dependent and ABA-independent pathways; whereas the cold stress response appears to be regulated essentially by ABA-independent mechanisms.Citation9 Stress inducible genes include rd (responsive to dehydration), erd (early responsive to dehydration), cor (cold regulated) and kin (cold-inducible) families in Arabidopsis.Citation10 Extensive transcriptional reprogramming under stress conditions is also evident in other plant species, although molecular studies on stress responses are dominated by Arabidopsis. For instance, several genes were reported to be differentially regulated in drought-tolerant and drought-sensitive genotypes of wild emmer wheat, the wild progenitor of modern bread wheat in response to slow and shock drought stresses.Citation11,Citation12 Interestingly, drought-tolerant genotype exhibited elevated levels of cold acclimation proteins even in the absence of stress and was also implicated to induce ABA-biosynthesis faster than the sensitive genotype in response to drought. However, both genotypes shared similar patterns of regulation for several genes, also present in Arabidopsis, suggesting that stress response mechanisms are essentially conserved among the plant kingdom with subtle differences among species and even genotypes that greatly affect the response outcome.Citation12
The cis-acting element, ABRE (ABA responsive element, PyACGTGG/TC), and its cognate bZIP family transcription factors (TFs) ABFs(ABRE binding factor)/AREBs(ABA responsive element binding protein) constitute one of the ABA-dependent pathways activated in response to particularly drought, salt and cold stresses.Citation1,Citation10,Citation13 The Arabidopsis genome encodes nine ABRE-binding protein homologs of group A bZIP TFs, eight of which (AREB1/ABF2, AREB2/ABF4, AREB3/DPBF3, ABF3/DPBF5, ABI5/DPBF1, EEL/DPBF4, DPBF2 and AT5G42910) are induced by drought and salt stresses, whereas the remaining ABF1 is induced by cold stress.Citation1,Citation9,Citation13 Recently, group S, C and G bZIP TFs involved in ABA-signaling were also discovered in soybean; these TFs are implicated in the regulation of erf5, kin1, cor15a, cor78a, p5cs1, dreb2a and cor47 expression. Usually, activation by ABA requires more than one ABRE in the responsive genes;Citation13 accordingly rd29b induced by drought and salinity via ABA-dependent pathways contains 2 ABREs in its promoter region.Citation10 In addition, ABRE-binding proteins AREB1, AREB2 and ABF3 have been implicated to form homo- or heterodimers inside the nucleus.Citation13 Induction by an ABRE-binding protein through the ABRE element has also been revealed to require coupling elements.Citation9 MYB/MYC TFs are another group of TFs involved in regulation of ABA-responsive stress genes. These TFs recognize a number of cis-elements and binding preferences have been attributed to the regulation of different sets of downstream genes. Indeed, overexpression of MYB76 was shown to upregulate expression of rd29b, dreb2a, p5cs, rd1, erd10 and cor78a, in constrast to MYB92 which upregulated the expression of dreb2a, p5cs, rd17, but downregulated expression of rd29b, cor78, cor6.6 and cor15a in soybean.Citation14 Interestingly, a single MYB TF may even trigger different responses in different plant species. For instance, Oryza sativa MYB4 leads to enhanced cold tolerance accompanied with a dwarf phenotype when overexpressed in Arabidopsis, enhanced tolerance against drought and viral diseases but not cold stress in tomato and enhanced drought and cold tolerance in apple.Citation9 Additionally MYC/MYB TFs may work together to produce a synergistic effect; among the downstream genes known to be activated cooperatively by MYC/MYB TFs is Arabidopsis rd22 gene to enhance the stress response.Citation3,Citation10,Citation15
On the other hand, C-repeat binding factor (CBF)/Dehydration responsive element binding (DREB) TFs, which belong to the AP2/ERF superfamily, constitute a class of plant specific TFs that relay stress signals in an ABA-independent manner. DREB TFs commonly recognize the DRE/CRT cis-acting element (G/ACCGAC) to activate stress responsive genes, including genes encoding LEA and HSP proteins, and are essentially subdivided into two groups: DREB1/CBF and DREB2, involved in cold stress and dehydration stress responses respectively.Citation1,Citation3,Citation9 The DREB1/CBF regulon has been shown to involve more than 40 genes all of which contain the DRE/CRT element in their promoter region and initiate extensive transcriptional reprogramming in response to cold stress. Inducer of cbf expression region 1 and 2 (ICEr1 and ICEr2) are recognized by regulators, such as the basic helix-loop-helix TF ICE1, to activate the DREB1/CBF regulon. Interestingly, MYB15 is also capable of driving the expression of CBF3 via Myb sequences located in the upstream of cbf3, similar to the activation of cbf2 by calmodulin-binding transcriptional activators (CAMTAs), suggesting that a complex network of regulators provides cross-talk between these stress response pathways. The second group of AP2/ERF family TFs, DREB2 proteins, is primarily induced by drought and salinity stresses. The heat shock transcription factor 3 in Arabidopsis (AtHSFA3) involved in thermo-tolerance was revealed to be a direct target of DREB2A, thereby linking DREB2 proteins to heat stress responses, as well.Citation1 Although stress responsive proteins are generally allocated to one or a few types of stress responses, it should be kept in mind that stress responses tend to be intermingled and individual components are strongly induced by one or a few of several stimuli.
Members of the NAM ATAF1 CUC2 (NAC) family of TFs are also generally regarded as ABA-independent stress response regulators. NAC proteins recognize and bind NAC recognition sites (NACRS) as multimers to activate gene expression.Citation5 One member of the NAC family, ERD1, participates in response mechanisms against drought, salinity and dark-induced senescence in an ABA-independent manner. In contrast, RD26, another member, has been shown to be induced both by drought and salinity, and by ABA. Furthermore, NAC family of transcription factors do not always enhance plant tolerance to stress conditions. For instance, ATAF1 is a negative regulator of the drought stress response, downregulating expression of drought-responsive cor47/rd17, erd10, kin1, rd22 and cor78. Consequently, ataf1 mutants recover better than wildtype plants upon drought.Citation9
Despite the large body of knowledge about the downstream effects of ABA-dependent stress-signaling, elucidation of the complete signal pathway was revealed only recently with the discovery of a small protein family that might act as ABA receptors and two putative ABA-transporters, both belonging to ATP-binding cassette (ABC) transporter family.Citation1,Citation8 PYR1/PYLs/RCARs (pyrabactin resistance 1/Pyr-likes/regulatory component of ABA receptors) were identified as proteins that can bind ABA and also interact with type 2C protein phosphatases (PP2C) such as ABA Insensitive 1 and 2 (ABI1 and ABI2), thus emerging as a putative receptor for ABA perception.Citation1 PYR1/RCARs are small soluble proteins from START/Bet V 1 superfamily. A hydrophobic pocket at the center binds ABA. In the absence of ABA, two loops surrounding the pocket assume an open-lid conformation. Upon binding, ABA triggers conformational changes which lock the ABA inside and expose a docking site at the surface of the closed lid. This docking site binds a PP2C via a conserved tryptophan residue and blocks the active site of the enzyme, inhibiting its phosphatase activity.Citation1,Citation4,Citation8 In the current model, SnRK2 kinases are dephosphorylated by PP2Cs in the absence of ABA, blocking downstream signaling. Upon ABA accumulation, PYRs/RCARs bind to it and the ABA-bound receptors inactivate PP2Cs, eliminating the inhibitory effect on SnRK2 kinases. SnRK2 kinases switch to the phosphorylated state, thereby activating bZIP family of TFs such as ABFs/AREB, and in turn activating downstream ABA-responsive genes.Citation1
The SnRK superfamily in plants consists of three sub-families of which SnRK1 subfamily members are orthologs of Saccharomyces cerevisiae Sucrose Non-fermenting 1 (SNF1) and mammalian AMP-activated protein kinase (AMPK). Plant SnRK1 kinases share approximately 62% and 48% of amino acid identity in the catalytic domain and overall, respectively, with SNF1 and AMPK kinases. In fact, all three protein families are so similar that peptides such as SAMS and AMARA are recognized by all three, enabling catalytic activity comparison.Citation16,Citation17 Recently, two SnRK1 family members KIN10 and KIN11 were implicated to initiate extensive transcriptional reprogramming in response to several stress signals. Particularly, KIN10 appears to regulate the expression of genes encoding histone deacetylases, signal transduction kinases and phosphatases and calcium modulators, in addition to several TFs.Citation18
It is noteworthy that the SnRK superfamily in plants is highly divergent. Whereas the Arabidopsis genome encodes three members of SnRK1 family, the barley genome encodes 10–20 which are further subdivided into SnRK1a and SnRK1b, the latter of which is exclusively found in cereals. Other members of the superfamily, SnRK2 and SnRK3 exhibit even more divergence, sharing only 42–45% amino acid identity in the catalytic domain with SnRK1s, SNF1s and AMPKs. Ten SnRK2s and 29 SnRK3s have been identified in Arabidopsis of which some members are involved in stress responses.Citation16,Citation17 For instance, nine of the Arabidopsis SnRK2s have been reported to be activated upon osmotic stress in a Ca2+-independent fashion, five have been shown to be induced by ABA treatment.Citation19 Additionally, PKABA1, which belongs to the SnRK2 family, is implicated in ABA-dependent regulation of gene expression.Citation16 Another well-known example is the transcriptional activation of salt overly sensitive 2 (SOS2) of the SnRK3 family due to salinity stress, where activated SOS2 complexes with SOS3 to activate a Na+/H+ antiporter, SOS1, at the same time upregulating its expression, eventually to maintain ion homeostasis.Citation19
Yeast SNF1 and mammalian AMPK are known to be induced by glucose starvation accompanied by a high AMP/ATP ratio.Citation16-Citation18 Energy status fluctuations are common in plants and can occur naturally whenever photosynthetic rate is affected due to limited access to sunlight, extension of night hours or in etiolated leaves of the seedlings, in addition to as a direct consequence of the competition among primary and secondary sink organs over photoassimilates.Citation18,Citation20 Direct activation of plant SnRK1 by sugars or AMP molecules is currently a controversial issue, although an indirect inhibitory effect of AMP on dephosphorylation of SnRK1 has been demonstrated.Citation21 However, overall energy status, which is closely linked to the sugar content of the cell, does appear to regulate SnRK1. In addition, glucose-6-phosphate, generated by hexokinase (HXK) during glucose metabolism, is capable of inhibiting SnRK1 activity.Citation22 Considering the dual function of HXK, for glycolysis in mitochondria and signaling in the nucleus, it has been proposed that HXK and SnRK1 pathways may compete to promote or arrest growth based on glucose availability. Consequently, energy signaling is now accepted to be linked to stress signaling. Furthermore, taking nitrogen, which affects the expression of many sugar-related genes, into account, it is plausible that nitrogen status could also be a determinant in the cross-talk between energy and stress signaling.Citation18
Mammalian AMPK is a negative regulator of the central protein kinase, target of rapamycin (TOR), responsible for cell growth and proliferation.Citation18 Similarly, plant TOR, which promotes cell growth under favorable conditions, when disrupted has been shown to cause early senescence in leaves, accumulation of soluble sugars, elevated levels of glutamine synthetase and glutamate dehydrogenase and early arrest of embryo development in Arabidopsis. In addition, Arabidopsis TOR expression was negatively affected by ABA accumulation and osmotic stress, whereas its overexpression could restore root growth to an extent in salinity stress conditions. Interestingly, the AtTOR-silenced phenotype in plantlets was reported to resemble the phenotype caused by overexpression of ABA-insensitive 5 (ABI5), a negative regulator of early events of ABA signaling,Citation23 suggesting a putative link between TOR regulated growth and ABA-signaling.
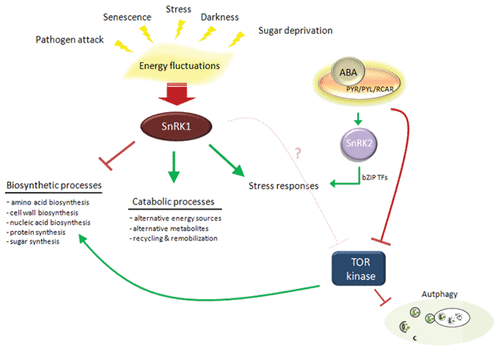
Although the interaction between mammalian AMPK and mTOR has been clearly demonstrated via competitive phosphorylation of Ulk1, homolog of autophagy initiator ATG1Citation22,Citation24 in addition to direct inhibition of mTOR by AMPK-mediated phosphorylation of an mTOR interacting protein,Citation22,Citation25 such an interaction between plant SnRK1 and TOR is not yet concrete.Citation26 However, given the evolutionary conservation of SnRK1/SNF1/AMPK and TOR signaling pathways, including the upstream kinases in plants, fungi and animals,Citation27 cross-talk between these pathways in plants would be plausible. In addition, the involvement of SnRK1 at the junction of sugar and ABA signaling at the onset of autotrophism in Arabidopsis suggests that SnRK1 may be a central regulator of metabolism, linking several anabolic and catabolic processes (Fig. 1).Citation28 On the other hand, since the expression of TOR is more prominent in meristematic tissues in plants, regulation of autophagy in mature plant cells was considered to be less likely to proceed via TOR.Citation29
Adverse conditions, such as nutrient deprivation and senescence, are known to enhance autophagy – a constitutive process where cytoplasmic constituents and particularly proteins are enclosed in double membrane vesicles called autophagosomes, degraded and recycled in bulk. Besides recycling and remobilization of nutrients, autophagy is also implicated in the turnover of long-lived proteins, and the removal of protein aggregates or damaged organelles which may otherwise cause disturbances. Consequently, autophagy is essential for normal plant growth processes, such as germination, seed development, hormone responses and pathogen resistance.Citation30,Citation31 Recently, plant TOR has been revealed as a negative regulator of autophagy, where reduced expression of TOR resulted in the enhancement of autophagy in an Atg18a-dependent manner in Arabidopsis.Citation32 Research on TOR signaling in plants is hampered compared with fungi and animals due to the embryonic lethality of TOR-knockout mutant plantsCitation22,Citation26; however, Liu and Bassham have successfully applied RNA interference to investigate effects of TOR on autophagy.Citation32
In plants, micro- and macroautophagy are well documented among the various types of autophagy, although a cytoplasm-to-vacuole route is also likely to exist.Citation33-Citation35 Macroautophagy, in which autophagic cargo is first encapsulated inside a double-membrane structure called ‘autophagosome’ and then directed to the vacuole for degradation, is essentially referred as ‘autophagy’. In addition, the eventual fusion of autophagosome with the vacuole has been proposed to follow at least two different routes in plants, either directly or via an intermediary fusion with an endosome-like organelle.Citation33 Hence, the contents of autophagosomes may well be degraded prior to fusion with the vacuole.Citation36 Under nutrient-rich conditions, TOR blocks the activation of autophagy through hyperphosphorylation of ATG1 thereby preventing its association with ATG13 at the earliest step of autophagosome formation. Starvation inhibits the growth-promoting activity of TOR, allowing ATG1 and ATG13 to assemble into an active complex with several other proteins to initiate autophagy. This active complex leads to two ubiquitin-like conjugation events that drive vesicle expansion and formation of enclosed autophagosomes. In plants, these two ubiquitin-like conjugation events comprise the most extensively studied parts of the molecular machinery of autophagy and conjugate ATG8 and ATG12 to phosphatidylethanolamine (PE) and ATG5, respectively. The Atg8-PE conjugate is anchored to the autophagosome membrane, whereas the ATG12-ATG5 conjugate is loosely bound by the autophagosome. Several other proteins direct the mature autophagosome to the vacuole, where the autophagosome contents are degraded by the actions of a variety of degrading enzymes, including hydrolases, proteases, nucleases and glucanases.Citation36,Citation37 Molecular events governing autophagy have been extensively studied in yeast, and the autophagy genes required for the core machinery were first isolated in yeast. Although several of these core genes are not yet identified or functionally characterized in animals, and to a greater extent in plants, progress so far suggests an evolutionarily conserved mechanism. Indeed, autophagy has been shown to be upregulated in response to abiotic stresses in even brown alga.Citation38 The availability of the whole genome sequence, in Arabidopsis, enabled the identification of more than 30 genes involved in autophagy.Citation39,Citation40 Recently, autophagy related genes have been identified in other organisms as well, such as rice, maize and wheat.Citation35,Citation41,Citation42 Excellent reviews on plant autophagy can be found elsewhere in the literature.Citation26,Citation33,Citation36,Citation37,Citation40,Citation43,Citation44
Autophagy has been implicated to play roles in a variety of abiotic and biotic stresses in plants; high salinity, drought, nutrient deprivation, reactive oxygen species and pathogen attack are all known to induce autophagy.Citation39,Citation45 Osmotic stress and salinity, among the most prevalent environmental stress factors, have recently been shown to activate autophagy and reactive oxygen species (ROS) generated in the process were proposed to act as potential signal transducers. Indeed, inhibition of NADPH oxidase activity mitigated autophagic response against salt but not osmotic stress, suggesting that both ROS-mediated and ROS-independent pathways to activate autophagy in response to stress conditions may exist.Citation30 In fact, ROS are generated in most if not all conditions compromising plant growth and may oxidize proteins and lipids and damage organelles if accumulated. Autophagy is often attributed to degrade these oxidized or damaged molecules in order to protect the cell.Citation33,Citation39,Citation46 Nutrient starvation, including carbon and nitrogen, is also known to elevate autophagic activity in plants, presumably to recycle and remobilize nutrients.Citation33,Citation39,Citation47 In C3 plants, up to 80% of leaf proteins are stored in chloroplasts, incorporated primarily into photosynthetic proteins, such as Rubisco. Therefore chloroplast proteins provide a rich source for nitrogen recycling when nitrogen is scarce. Chloroplast degradation via autophagy has recently emerged as a novel route to relocate nitrogen in starved plants, where Rubisco-containing bodies (RCBs), containing stromal proteins and chloroplast envelope, are degraded to release carbon and nitrogen for reuse.Citation48
Taking into account the intermingled signaling pathways of carbon and nitrogen metabolism, a recently emerged view indicates that autophagy may play another, potentially more important, role in stress responses other than nutrient recycling and degradation of stress-damaged proteins and organelles. Indeed, RNAi of Atg18a in Arabidopsis did not significantly elevated the oxidized protein levels compared with wildtype plants, leading to the conclusion that autophagy may not be central to the elimination of ROS-damaged proteins or protein aggregates.Citation30 Additionally, the recent discovery of a putative selective route for autophagic degradation through the interaction with autophagosome-membrane-bound ATG8 protein and proteins carrying an ATG8-interacting motif (AIM),Citation31 shows that besides bulk degradation, autophagy is also capable of regulating stress responses by selective degradation of signal transducers to modulate the response over time. Indeed, a stress-responsive membrane protein tryptophan-rich sensory protein related (TSPO) has been shown to be targeted by autophagy pathway via its AIM, and degraded.Citation49,Citation50 In a very recent study, upon ABA-perception, TSPO expression was shown to be upregulated with a concomitant increase in heme content in Arabidopsis. Heme molecules, as other porphyrins, may cause cytotoxicity by light-dependent generation of radicals when unbound by other proteins. On the other hand, several ROS-scavenging enzymes require porphyrin cofactors. Consequently, it has been proposed that ABA or other stress factors result in the generation of ROS that may damage cell components. To protect the cell from oxidative damage, ROS-scavenging enzymes are recruited, leading to an increase in the porphyrin levels, such as heme, as well. However, because porphyrins are cytotoxic, their levels are tightly regulated; a concomitant increase in TSPO captures any unbound heme and triggers its degradation via autophagy.Citation50
The recent discovery of a selective route of autophagic degradation supports the pivotal role of autophagy in plant growth and development, as well as in stress responses.Citation33,Citation51 The Arabidopsis genome encodes nine ATG8 protein homologs that function in autophagyCitation52 and, although ATG8 has long been considered as a hallmark of autophagy, it was only recently that ATG8 was attributed to play a central role in autophagy, particularly selective autophagy. Upon the identification of the ATG8-interacting domain, a large number of proteins have been shown to interact with ATG8. Consequently, ATG8 is now being speculated to act as a docking platform for selective cargo.Citation31 The majority of the plant studies investigating the role of autophagy genes have been dependent upon T-DNA insertion or stable RNAi constructs.Citation36,Citation37,Citation47 A number of studies have successfully applied Virus-Induced Gene Silencing (VIGS), as well, to downregulate gene expression.Citation35,Citation45 VIGS provides an efficient means of carrying out functional studies and may also be a viable alternative when the disruption of a gene of interest causes lethality and also in monocot species for which transformation methods are limited. Furthermore, autophagy proteins, particularly ATG8 and ATG18, are encoded by multigene families, rendering knockout studies unfeasible or labor-intensive due to functional redundancy.Citation40,Citation53 In contrast, an entire multigene family can be targeted, and all related transcripts can be downregulated by VIGS.Citation35 Functional studies of several autophagy-related genes, particularly Atg8, appear promising in understanding of the crosstalk between growth promoting and growth limiting scenarios in plants. In addition, identification of novel stress inducible genes and characterization of the respective transcripts may provide clues on missing points in our understanding of the complex responses of plants against multiple environmental stimuli. In one such study, a transmembrane protein, TdicTMPIT1, was shown to be differentially regulated in response to drought stress in wild emmer wheat but not in moden durum wheat. Sequence analysis of TdicTMPIT1 revealed significant similarity to TransMembrane Protein Inducible by TNF-α (TMPIT) family and its differential expression in response to drought suggested a putative role in stress signaling that is yet-to-be-elucidated.Citation54 Identification and characterization of such novel proteins may establish novel links between cooperating signaling pathways. Coupled with next generation genomics tools, such information will be valuable in the generation of stress-tolerant plants.
Figure 1. Sucrose Non-fermenting 1 (SNF1) related kinases (SnRKs) appear as central regulators of energy and stress signaling pathways in plants. Direct or indirect regulation of autophagy by this family of kinases remains to be elucidated.
Related Research Data
References
- Qin F, Shinozaki K, Yamaguchi-Shinozaki K. Achievements and challenges in understanding plant abiotic stress responses and tolerance. Plant Cell Physiol 2011; 52:1569 - 82; 10.1093/pcp/pcr106; PMID: 21828105
- Knight H, Knight MR. Abiotic stress signalling pathways: specificity and cross-talk. Trends Plant Sci 2001; 6:262 - 7; 10.1016/S1360-1385(01)01946-X; PMID: 11378468
- Hirayama T, Shinozaki K. Research on plant abiotic stress responses in the post-genome era: past, present and future. Plant J 2010; 61:1041 - 52; 10.1111/j.1365-313X.2010.04124.x; PMID: 20409277
- Raghavendra AS, Gonugunta VK, Christmann A, Grill E. ABA perception and signalling. Trends Plant Sci 2010; 15:395 - 401; 10.1016/j.tplants.2010.04.006; PMID: 20493758
- Nakashima K, Takasaki H, Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K. NAC transcription factors in plant abiotic stress responses. Biochim Biophys Acta 2012; 1819:97 - 103; 10.1016/j.bbagrm.2011.10.005; PMID: 22037288
- Finkelstein RR, Gampala SS, Rock CD. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002; 14:Suppl S15 - 45; PMID: 12045268
- Tuteja N. Abscisic Acid and abiotic stress signaling. Plant Signal Behav 2007; 2:135 - 8; 10.4161/psb.2.3.4156; PMID: 19516981
- Hubbard KE, Nishimura N, Hitomi K, Getzoff ED, Schroeder JI. Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions. Genes Dev 2010; 24:1695 - 708; 10.1101/gad.1953910; PMID: 20713515
- Agarwal PK, Jha B. Transcription factors in plants and ABA dependent and independent abiotic stress signalling. Biol Plant 2010; 54:201 - 12; 10.1007/s10535-010-0038-7
- Shinozaki K, Yamaguchi-Shinozaki K. Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr Opin Plant Biol 2000; 3:217 - 23; PMID: 10837265
- Ergen NZ, Budak H. Sequencing over 13 000 expressed sequence tags from six subtractive cDNA libraries of wild and modern wheats following slow drought stress. Plant Cell Environ 2009; 32:220 - 36; 10.1111/j.1365-3040.2008.01915.x; PMID: 19054353
- Ergen NZ, Thimmapuram J, Bohnert HJ, Budak H. Transcriptome pathways unique to dehydration tolerant relatives of modern wheat. Funct Integr Genomics 2009; 9:377 - 96; 10.1007/s10142-009-0123-1; PMID: 19330365
- Fujita Y, Fujita M, Shinozaki K, Yamaguchi-Shinozaki K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J Plant Res 2011; 124:509 - 25; 10.1007/s10265-011-0412-3; PMID: 21416314
- Liao Y, Zou HF, Wang HW, Zhang WK, Ma B, Zhang JS, et al. Soybean GmMYB76, GmMYB92, and GmMYB177 genes confer stress tolerance in transgenic Arabidopsis plants. Cell Res 2008; 18:1047 - 60; 10.1038/cr.2008.280; PMID: 18725908
- Abe H, Yamaguchi-Shinozaki K, Urao T, Iwasaki T, Hosokawa D, Shinozaki K. Role of arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 1997; 9:1859 - 68; PMID: 9368419
- Halford NG, Hey S, Jhurreea D, Laurie S, McKibbin RS, Zhang Y, et al. Highly conserved protein kinases involved in the regulation of carbon and amino acid metabolism. J Exp Bot 2004; 55:35 - 42; 10.1093/jxb/erh019; PMID: 14645392
- Halford NG, Hey S, Jhurreea D, Laurie S, McKibbin RS, Paul M, et al. Metabolic signalling and carbon partitioning: role of Snf1-related (SnRK1) protein kinase. J Exp Bot 2003; 54:467 - 75; 10.1093/jxb/erg038; PMID: 12508057
- Baena-González E, Sheen J. Convergent energy and stress signaling. Trends Plant Sci 2008; 13:474 - 82; 10.1016/j.tplants.2008.06.006; PMID: 18701338
- Boudsocq M, Barbier-Brygoo H, Laurière C. Identification of nine sucrose nonfermenting 1-related protein kinases 2 activated by hyperosmotic and saline stresses in Arabidopsis thaliana. J Biol Chem 2004; 279:41758 - 66; 10.1074/jbc.M405259200; PMID: 15292193
- Usadel B, Bläsing OE, Gibon Y, Retzlaff K, Höhne M, Günther M, et al. Global transcript levels respond to small changes of the carbon status during progressive exhaustion of carbohydrates in Arabidopsis rosettes. Plant Physiol 2008; 146:1834 - 61; 10.1104/pp.107.115592; PMID: 18305208
- Sugden C, Crawford RM, Halford NG, Hardie DG. Regulation of spinach SNF1-related (SnRK1) kinases by protein kinases and phosphatases is associated with phosphorylation of the T loop and is regulated by 5′-AMP. Plant J 1999; 19:433 - 9; 10.1046/j.1365-313X.1999.00532.x; PMID: 10504565
- Robaglia C, Thomas M, Meyer C. Sensing nutrient and energy status by SnRK1 and TOR kinases. Curr Opin Plant Biol 2012; 15:301 - 7; 10.1016/j.pbi.2012.01.012; PMID: 22305521
- Deprost D, Yao L, Sormani R, Moreau M, Leterreux G, Nicolaï M, et al. The Arabidopsis TOR kinase links plant growth, yield, stress resistance and mRNA translation. EMBO Rep 2007; 8:864 - 70; 10.1038/sj.embor.7401043; PMID: 17721444
- Kim J, Kundu M, Viollet B, Guan KL. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat Cell Biol 2011; 13:132 - 41; 10.1038/ncb2152; PMID: 21258367
- Gwinn DM, Shackelford DB, Egan DF, Mihaylova MM, Mery A, Vasquez DS, et al. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol Cell 2008; 30:214 - 26; 10.1016/j.molcel.2008.03.003; PMID: 18439900
- Reumann S, Voitsekhovskaja O, Lillo C. From signal transduction to autophagy of plant cell organelles: lessons from yeast and mammals and plant-specific features. Protoplasma 2010; 247:233 - 56; 10.1007/s00709-010-0190-0; PMID: 20734094
- Polge C, Thomas M. SNF1/AMPK/SnRK1 kinases, global regulators at the heart of energy control?. Trends Plant Sci 2007; 12:20 - 8; 10.1016/j.tplants.2006.11.005; PMID: 17166759
- Jossier M, Bouly JP, Meimoun P, Arjmand A, Lessard P, Hawley S, et al. SnRK1 (SNF1-related kinase 1) has a central role in sugar and ABA signalling in Arabidopsis thaliana. Plant J 2009; 59:316 - 28; 10.1111/j.1365-313X.2009.03871.x; PMID: 19302419
- Menand B, Desnos T, Nussaume L, Berger F, Bouchez D, Meyer C, et al. Expression and disruption of the Arabidopsis TOR (target of rapamycin) gene. Proc Natl Acad Sci U S A 2002; 99:6422 - 7; 10.1073/pnas.092141899; PMID: 11983923
- Liu YM, Xiong Y, Bassham DC. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy 2009; 5:954 - 63; 10.4161/auto.5.7.9290; PMID: 19587533
- Li F, Vierstra RD. Autophagy: a multifaceted intracellular system for bulk and selective recycling. Trends Plant Sci 2012; In Press 10.1016/j.tplants.2012.05.006; PMID: 22694835
- Liu Y, Bassham DC. TOR is a negative regulator of autophagy in Arabidopsis thaliana. PLoS One 2010; 5:e11883; 10.1371/journal.pone.0011883; PMID: 20686696
- Bassham DC. Plant autophagy--more than a starvation response. Curr Opin Plant Biol 2007; 10:587 - 93; 10.1016/j.pbi.2007.06.006; PMID: 17702643
- Thompson AR, Doelling JH, Suttangkakul A, Vierstra RD. Autophagic nutrient recycling in Arabidopsis directed by the ATG8 and ATG12 conjugation pathways. Plant Physiol 2005; 138:2097 - 110; 10.1104/pp.105.060673; PMID: 16040659
- Kuzuoglu-Ozturk D, Cebeci Yalcinkaya O, Akpinar BA, Mitou G, Korkmaz G, Gozuacik D, et al. Autophagy-related gene, TdAtg8, in wild emmer wheat plays a role in drought and osmotic stress response. Planta 2012; In Press 10.1007/s00425-012-1657-3; PMID: 22569921
- Bassham DC, Laporte M, Marty F, Moriyasu Y, Ohsumi Y, Olsen LJ, et al. Autophagy in development and stress responses of plants. Autophagy 2006; 2:2 - 11; PMID: 16874030
- Thompson AR, Vierstra RD. Autophagic recycling: lessons from yeast help define the process in plants. Curr Opin Plant Biol 2005; 8:165 - 73; 10.1016/j.pbi.2005.01.013; PMID: 15752997
- Dittami SM, Scornet D, Petit JL, Ségurens B, Da Silva C, Corre E, et al. Global expression analysis of the brown alga Ectocarpus siliculosus (Phaeophyceae) reveals large-scale reprogramming of the transcriptome in response to abiotic stress. Genome Biol 2009; 10:R66; 10.1186/gb-2009-10-6-r66; PMID: 19531237
- Han S, Yu B, Wang Y, Liu Y. Role of plant autophagy in stress response. Protein Cell 2011; 2:784 - 91; 10.1007/s13238-011-1104-4; PMID: 22058033
- Avin-Wittenberg T, Honig A, Galili G. Variations on a theme: plant autophagy in comparison to yeast and mammals. Protoplasma 2012; 249:285 - 99; 10.1007/s00709-011-0296-z; PMID: 21660427
- Chung T, Suttangkakul A, Vierstra RD. The ATG autophagic conjugation system in maize: ATG transcripts and abundance of the ATG8-lipid adduct are regulated by development and nutrient availability. Plant Physiol 2009; 149:220 - 34; 10.1104/pp.108.126714; PMID: 18790996
- Xia K, Liu T, Ouyang J, Wang R, Fan T, Zhang M. Genome-wide identification, classification, and expression analysis of autophagy-associated gene homologues in rice (Oryza sativa L.). DNA Res 2011; 18:363 - 77; 10.1093/dnares/dsr024; PMID: 21795261
- Bassham DC. Function and regulation of macroautophagy in plants. Biochim Biophys Acta 2009; 1793:1397 - 403; 10.1016/j.bbamcr.2009.01.001; PMID: 19272302
- Il Kwon S, Park OK. Autophagy in Plants. J Plant Biol 2008; 51:313 - 20; 10.1007/BF03036132
- Liu Y, Schiff M, Czymmek K, Tallóczy Z, Levine B, Dinesh-Kumar SP. Autophagy regulates programmed cell death during the plant innate immune response. Cell 2005; 121:567 - 77; 10.1016/j.cell.2005.03.007; PMID: 15907470
- Xiong Y, Contento AL, Nguyen PQ, Bassham DC. Degradation of oxidized proteins by autophagy during oxidative stress in Arabidopsis. Plant Physiol 2007; 143:291 - 9; 10.1104/pp.106.092106; PMID: 17098847
- Yoshimoto K. Beginning to understand autophagy, an intracellular self-degradation system in plants. Plant Cell Physiol 2012; 53:1355 - 65; 10.1093/pcp/pcs099; PMID: 22764279
- Izumi M, Wada S, Makino A, Ishida H. The autophagic degradation of chloroplasts via rubisco-containing bodies is specifically linked to leaf carbon status but not nitrogen status in Arabidopsis. Plant Physiol 2010; 154:1196 - 209; 10.1104/pp.110.158519; PMID: 20807997
- Vanhee C, Batoko H. Autophagy involvement in responses to abscisic acid by plant cells. Autophagy 2011; 7:655 - 6; 10.4161/auto.7.6.15307; PMID: 21460617
- Vanhee C, Zapotoczny G, Masquelier D, Ghislain M, Batoko H. The Arabidopsis multistress regulator TSPO is a heme binding membrane protein and a potential scavenger of porphyrins via an autophagy-dependent degradation mechanism. Plant Cell 2011; 23:785 - 805; 10.1105/tpc.110.081570; PMID: 21317376
- Mitou G, Budak H, Gozuacik D. Techniques to study autophagy in plants. Int J Plant Genomics 2009; 2009:451357; 10.1155/2009/451357; PMID: 19730746
- Yoshimoto K, Hanaoka H, Sato S, Kato T, Tabata S, Noda T, et al. Processing of ATG8s, ubiquitin-like proteins, and their deconjugation by ATG4s are essential for plant autophagy. Plant Cell 2004; 16:2967 - 83; 10.1105/tpc.104.025395; PMID: 15494556
- Slavikova S, Ufaz S, Avin-Wittenberg T, Levanony H, Galili G. An autophagy-associated Atg8 protein is involved in the responses of Arabidopsis seedlings to hormonal controls and abiotic stresses. J Exp Bot 2008; 59:4029 - 43; 10.1093/jxb/ern244; PMID: 18836138
- Lucas S, Dogan E, Budak H. TMPIT1 from wild emmer wheat: first characterisation of a stress-inducible integral membrane protein. Gene 2011; 483:22 - 8; 10.1016/j.gene.2011.05.003; PMID: 21635942