Abstract
Plant-parasitic cyst nematodes form a specialized feeding site, termed a syncytium, in the roots of host plants. Monoclonal antibodies to defined glycans, in addition to a cellulose-binding module, were used to characterize the cell walls of a functioning syncytia in situ. Cell walls of syncytia were found to contain cellulose, xyloglucan and mannan. Analysis of the pectin network revealed syncytial cell walls are abundant in homogalacturonan, which was heavily methyl-esterified. Arabinan was also detected and the results suggest the cell walls of syncytia are highly flexible.
Plant-parasitic cyst nematodes, such as Heterodera schachtii, form a specialized feeding site in the roots of host plants through which the nematode obtains plant-derived nutrients. The feeding site, termed a syncytium, is formed from a single vascular cylinder root cell, generally a procambial or pericycle cell (for review see ref. Citation1). The development of syncytia is associated with proliferation of procambial cells that are subsequently incorporated into the growing feeding site through extensive cell wall remodelling and cell wall degradation processes.Citation1 Many previous studies have focused on elucidating the cell wall remodelling processes that occur during syncytial development.Citation2-Citation4 However, little was known about the structural architecture of syncytial cell walls. We recently used monoclonal antibodies to defined glycans and a cellulose-binding module to characterize the cell walls of syncytia formed by H. schachtii developing in A. thaliana.Citation5 Transverse sections were taken through syncytia at 14 d post inoculation (dpi) and the cell wall architecture of syncytia and surrounding stele cells was characterized in situ.
Fluorescent imaging was used to analyze cellulose, hemicelluloses (xyloglucan and mannan) and pectin (homogalacturonan and rhamnogalacturonan-I), which constitute the carbohydrate component of plant cell walls. Additionally, the cell wall structural proteins arabinogalactan proteins (AGPs) and extensins were investigated. The immunolabelling of cellulose, xyloglucan and mannan (using CBM3a; LM15 and LM21antibodies respectively) were widely similar in sections taken through nematode infected root sections in comparison to uninfected roots. Cellulose was detected in cell walls of the syncytia () along with xyloglucan and mannan. It has been proposed that the hemicelluloses form cross-linkages with the cellulose microfibrils and provide the load-bearing structure in the syncytial cell wall.Citation6
Figure 1. Fluorescent imaging of syncytial cell wall components. (A) Cellulose-binding module 3a (CBM3a) was used to visualize crystalline cellulose in the cell walls of syncytia formed by H. schachtii in A. thaliana at 14 dpi. (B) Transverse sections through nematode infected roots were pre-treated with 0.1 M sodium carbonate to remove methyl ester groups from pectin HG chains. Following pre-treatment the sections were treated with the LM19 antibody, which bound to the cell walls across the stele. (C) Immunolabelling with the LM20 antibody, which binds to methyl-esterified pectin HG resulted in strong fluorescence in all cell walls. Scale bars, 50 µm.
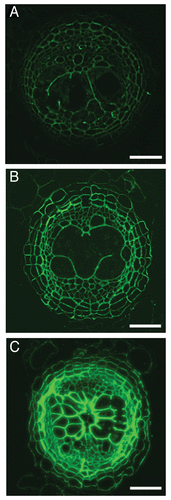
A range of monoclonal antibodies were used to characterize the pectin network of syncytial cell walls (). Pectin refers to a group of complex polysaccharides which includes homogalacturonan (HG) and rhamnogalacturonan-I (RG-I). The antibodies LM19 and LM20 were used to localize pectin HG in nematode infected root sections and determine the methyl-esterification status of the polysaccharide. The LM19 antibody preferentially binds to de-esterified pectin HGCitation7 and in nematode infected root sections the epitope was detected in pericycle cells only. However, immunolabelling with the LM19 antibody following the pre-treatment of root sections with 0.1 M sodium carbonate, which removes methyl esters from HG, resulted in strong binding of the antibody in all cells within the stele of nematode infected root sections (with the exception of xylem vessels; ). This was confirmed with immunolabelling of the LM20 antibody, which binds to methyl-esterified pectin HG.Citation7 The LM20 antibody bound extremely strongly across the stele in nematode infected root sections (). In uninfected root sections the LM20 epitope was detected at a low level. The LM19 and LM20 antibody immunolabelling collectively demonstrate that syncytial cell walls are abundant in pectin HG, which is heavily methyl-esterified. The pectin network is a major component of plant cell walls and the methyl-esterification status of pectin HG influences the mechanical properties and porosity of cell walls.Citation8 There is a body of evidence that suggests homogalacturonan is synthesized in the Golgi apparatus where 70–80% of the galacturonic acid residues are methyl-esterified prior to secretion into the primary cell wall (for review seeCitation9). Methyl-ester groups are subsequently removed from regions of HG in the cell wall by the enzymatic action of pectin methylesterases.Citation10 De-esterified regions of HG can form cross-linkages with calcium ions, which can reduce porosity and strengthen the cell wall.Citation11,Citation12 The heavily methyl-esterified status of the pectin HG in the cell walls of syncytia is predicted to result in a highly flexible wall. Flexibility is an important property of syncytial cell walls as the cell must withstand high turgor pressures, which arise from solutes accumulating in the syncytia.Citation13 Additionally, the feeding site must maintain structural integrity following the nematode ingesting the cytoplasmic contents of the syncytium.
Table 1. Overview of monoclonal antibodies used to characterize the structural architecture of syncytial cell walls together with details of antibody epitope detection.
While the highly methyl-esterified status of pectin HG provides an obvious function in terms of cell wall flexibility, it does raise questions with regards to cell wall degradation processes. For cell wall degradation to occur, the disassembly of all cell wall polymers is required. Pectin chains are hydrolysed by pectate lyases and polygalacturonase enzymes however, these enzymes are unable to act upon pectin HG which is methyl esterified (for review seeCitation14). During fruit ripening, a process that requires extensive cell wall degradation, the de-esterification of methyl-esterified HG is an important step in the degradation process.Citation15-Citation17 Therefore, at 14 dpi the syncytium is a cell in which extensive cell wall degradation has occurred but pectin, one of the major polysaccharide components of the cell wall, is in a condition that is protected from degradation. We characterized syncytia the feeding size had reached the maximum size, as a result we revealed the cell wall structural architectural requirements for syncytial function as opposed to formation. We will now go on to determine the methyl-esterification status of pectin HG during the early stages of syncytial development, when the extensive cell wall remodelling and degradation processes are occurring. This will hopefully provide an insight into how the dynamic cell wall adapts to these conflicting requirements.
| Abbreviations: | ||
| dpi | = | days post inoculation |
| HG | = | homogalacturonan |
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Related Research Data
References
- Sobczak M, Golinowski W. Cyst nematodes and syncytia. In: Jones J, Gheysen G, Fenoll C, eds. Genomics and Molecular Genetics of Plant-Nematode Interactions. Berlin: Springer, 2011:61-82.
- Wieczorek K, Golecki B, Gerdes L, Heinen P, Szakasits D, Durachko DM, et al. Expansins are involved in the formation of nematode-induced syncytia in roots of Arabidopsis thaliana.. Plant J 2006; 48:98 - 112; 10.1111/j.1365-313X.2006.02856.x; PMID: 16942607
- Wieczorek K, Hofmann J, Blöchl A, Szakasits D, Bohlmann H, Grundler FMW. Arabidopsis endo-1,4-beta-glucanases are involved in the formation of root syncytia induced by Heterodera schachtii.. Plant J 2008; 53:336 - 51; 10.1111/j.1365-313X.2007.03340.x; PMID: 18069944
- Sobczak M, Fudali S, Wieczorek K. Cell wall modifications induced by nematodes. In: Jones J, Gheysen G, Fenoll C, eds. Genomics and Molecular Genetics of Plant-Nematode Interactions. Berlin: Springer, 2011:395 - 423.
- Davies LJ, Lilley CJ, Paul Knox J, Urwin PE. Syncytia formed by adult female Heterodera schachtii in Arabidopsis thaliana roots have a distinct cell wall molecular architecture. New Phytol 2012; 196:238 - 46; 10.1111/j.1469-8137.2012.04238.x; PMID: 22803660
- Cosgrove DJ. Growth of the plant cell wall. Nat Rev Mol Cell Biol 2005; 6:850 - 61; 10.1038/nrm1746; PMID: 16261190
- Verhertbruggen Y, Marcus SE, Haeger A, Ordaz-Ortiz JJ, Knox JP. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr Res 2009; 344:1858 - 62; 10.1016/j.carres.2008.11.010; PMID: 19144326
- Willats WGT, Orfila C, Limberg G, Buchholt HC, van Alebeek GJ, Voragen AGJ, et al. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls. Implications for pectin methyl esterase action, matrix properties, and cell adhesion. J Biol Chem 2001; 276:19404 - 13; 10.1074/jbc.M011242200; PMID: 11278866
- Mohnen D. Pectin structure and biosynthesis. Curr Opin Plant Biol 2008; 11:266 - 77; 10.1016/j.pbi.2008.03.006; PMID: 18486536
- Micheli F. Pectin methylesterases: cell wall enzymes with important roles in plant physiology. Trends Plant Sci 2001; 6:414 - 9; 10.1016/S1360-1385(01)02045-3; PMID: 11544130
- Willats WGT, McCartney L, Mackie W, Knox JP. Pectin: cell biology and prospects for functional analysis. Plant Mol Biol 2001; 47:9 - 27; 10.1023/A:1010662911148; PMID: 11554482
- Derbyshire P, McCann MC, Roberts K. Restricted cell elongation in Arabidopsis hypocotyls is associated with a reduced average pectin esterification level. BMC Plant Biol 2007; •••:7; PMID: 17302987
- Böckenhoff A, Prior DAM, Grundler FMW, Oparka KJ. Induction of phloem unloading in Arabidopsis thaliana roots by the parasitic nematode Heterodera schachtii.. Plant Physiol 1996; 112:1421 - 7; 10.1104/pp.112.4.1421; PMID: 8972592
- Wolf S, Mouille G, Pelloux J. Homogalacturonan methyl-esterification and plant development. Mol Plant 2009; 2:851 - 60; 10.1093/mp/ssp066; PMID: 19825662
- Roy S, Vian B, Roland JC. Immunocytochemical study of the deesterification patterns during cell-wall autolysis in the ripening of cherry tomato. Plant Physiol Biochem 1992; 30:139 - 46
- Blumer JM, Clay RP, Bergmann CW, Albersheim P, Darvill A. Characterization of changes in pectin methylesterase expression and pectin esterification during tomato fruit ripening. Can J Bot 2000; 78:607 - 18
- Brummell DA, Harpster MH. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol Biol 2001; 47:311 - 40; 10.1023/A:1010656104304; PMID: 11554479