Abstract
MicroRNAs (miRNAs) are a class of small non-coding RNAs (small RNAs) that are 20–24nt in length and predominantly repress gene expression at post-transcriptional levels. They regulate many biological processes including development, metabolism and physiology. Numerous studies have revealed that the steady-state levels of miRNA are under sophisticated control to ensure their proper function. In this review, we summarize recent advances on regulation of miRNA processing and stability in plants.
Plants use multiple mechanisms to refine gene expression in order to maintain their normal development and physiology. miRNAs are repressors of gene expression in plants. They associate with and guide the Agronaute (AGO) protein to cleave and/or inhibit translation of mRNAs containing their complementary sequences. Since first isolated in 2002,Citation1-Citation3 hundreds of plant miRNAs have been identified. Some miRNAs are highly conserved among different plant species.Citation4 These miRNAs regulate the expression of hundreds of genes, most of which are transcription factors involved in various biological processes such as development, metabolism and plant responses to biotic and abiotic stresses.Citation5,Citation6
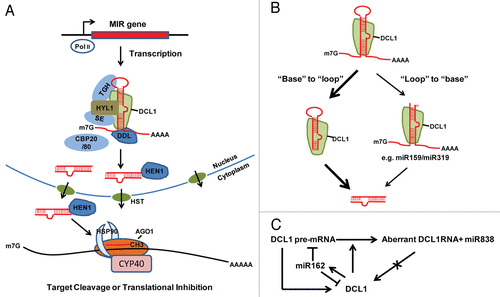
miRNAs are derived from primary transcripts called pri-miRNAs, which are encoded by miRNA genes. Many miRNA genes are located in intergenic regions while a few reside in the introns of host genes. Like mRNAs, pri-miRNAs are mainly transcribed by RNA polymerase II, which are followed by 5′ capping, 3′ polyadenylation and/or splicing.Citation7,Citation8 A characteristic feature of pri-miRNAs is the presence of imperfect stem-loop structure with long flanking arms (also termed hairpin structure), which can be recognized by RNase III family enzymes.Citation9 The framework of plant miRNA biogenesis has been established in model plant Arabidopsis thaliana (). In Arabidopsis, RNase III enzyme DICER-LIKE 1 (DCL1) releases miRNA duplex from pri-miRNAs. The miRNA duplex contains a miRNA strand and a passenger strand called miRNA*.Citation10 DCL4, a homolog of DCL1, is responsible for the generation of some miRNAs as well.Citation11 After production, miRNAs are loaded into AGO1 to form an active RNA-induced silencing complex (RISC), while the miRNA* is degraded.Citation12-Citation14 RISC formation is an ATP-dependent process and requires CYCLOPHILIN 40 (CYP40) and HEAT SHOCK PROTEIN 90 (HSP90)Citation15-Citation17 (). Studies have established that miRNA levels depend on the combination effect of biogenesis, RISC loading and degradation. Due to space limitation, we focus on the regulatory mechanisms on pri-miRNA processing and miRNA turn over in this mini review.
Figure 1. The miRNA pathway in Arabidopsis. A. miRNA biogenesis. MIR genes are transcribed by Pol II. Like mRNAs, primary transcripts of miRNAs (pri-miRNAs) are 5′ capped and 3′ polyadenylated. The DCL1 complex recognizes hairpin structure of pri-miRNA and catalyzes sequential cleavage to release miRNA/miRNA* duplex. The duplex is then exported to the cytoplasm by HASTY (HST) and loaded into an AGO1 complex to form a functional RISC complex. HEN1 prevents small RNA degradation by the addition of a methyl group to the last nucleotide of miRNA/miRNA* duplex. B. Two ways of pri-miRNA processing in plants. Most pri-miRNAs use a conical “base” to “loop” processing pathway. Instead, a few pri-miRNAs containing unusual long stem structure may employ a nonconical “loop” to “base” processing pathway. C. Autoregulation and feedback regulation of DCL1. DCL1 is required for miR162 maturation, which in turn guides DCL1 mRNA cleavage to repress DCL1 expression. In addition, DCL1 acts on its own pre-mRNA to produce miR838, resulting in aberrant DCL1 RNA that fails to produce functional DCL1.
Regulation of Pri-miRNA Processing
DCL1 forms a complex with the dsRNA-binding protein HYPONASTIC LEAVES1 (HYL1) and zinc-finger protein SERRATE (SE) to process pri-miRNAs.Citation18-Citation20 HYL1 and SE are required for efficient and accurate processing of pri-miRNAs.Citation21-Citation25 A majority of pri-miRNAs are first cleaved by the DCL1 complex to release the stem-loop precursors (pre-miRNAs), which are further processed by DCL1 to generate the miRNA duplexes ().Citation26,Citation27 This conventional “base” to “loop” processing is similar to those in animals.Citation28 Studies on several miRNAs reveal that a double strand to single strand transition at 14–17 nt proximal to the mature miRNA:miRNA* duplex is essential for the accurate processing.Citation26,Citation27,Citation29,Citation30 This stem segment is believed to be the docking site for DCL1 to execute a first cut, and then a second cut is made by measuring 21nt from the first cut. However, pri-miR159 and pri-miR319 are not processed from “base” to “loop”Citation31,Citation32 (). Their processing starts with a cut just below the terminal loop, which is followed by three additional cleavages.Citation31,Citation32 This variation may be caused by their long upper stem structure.Citation32 In Arabidopsis, like pri-miR159 and pri-miR319, 17 additional pri-miRNAs contain long stem loop structure and generate two or more miRNAs from the same precursor.Citation33 It is possible that these precursors may also utilize this non-conical mechanism.
Analysis of pri-miRNA levels using dcl1, hyl1 and se mutants implicates tissue- and precursor-specific patterning of pri-miRNA processing,Citation34 indicating that the activity of DCL1 complex is regulated.Citation34 The core component, DCL1 itself is under stringent feedback regulation by one of its product, miR162Citation35 (). In addition, DCL1 can act on its own pre-mRNA to release pre-miR838, which produces nonfunctional DCL1 pre-mRNA and therefore, regulates the levels of DCL1Citation11 (). The Short Interspaced Elements-Derived RNAs (SINE RNAs) have been shown to control the HYL1 activity.Citation36 Because SINE RNAs are structurally similar to pri-miRNAs, they will compete with pri-miRNAs for HYL1 and therefore modulate the processing of pri-miRNAs through tissue- or developmental stage-specific expression.Citation36
The activity of DCL1 is also controlled by factors other than HYL1 and SE. Gel filtration analysis revealed that the DCL1 complex is about 660kDa in size, suggestive of the existence of additional components.Citation13 Indeed, the RNA-binding proteins DAWDLE (DDL) and TOUGH (TGH) are identified as DCL1 interacting proteinsCitation37,Citation38 (). DDL is a forkhead-associated domain-containing protein and is required for the accumulation of miRNAs.Citation38 DDL interacts with DCL1 and binds pri-miRNA162b in vitro, supporting its role in facilitating pri-miRNA processing. However, DDL seems to have additional roles in the miRNA pathway, because pri-miRNA levels are reduced in ddl while they are increased in dcl1. However, DDL does not affect the transcription of MIR genes, suggesting that DDL may stabilize pri-miRNAs. TGH contains a G-patch and a SWAP domain in its N-terminus, which are often found in RNA binding and/or processing related proteins.Citation39 TGH binds both pri-miRNAs and pre-miRNAs and interacts with DCL1, HYL1 and SE.Citation37 Mutation of TGH impairs DCL1 activities in vitro and reduces the accumulation of miRNAs.Citation37 In addition, tgh-1 reduces the amount of pri-miRNAs in the DCL1 complex.Citation37 These facts suggest that TGH may promote DCL1 activity and facilitate the recruitment of pri-miRNAs to the DCL1 complex. Unlike SE and HYL1, TGH may not contribute to the DCL1 processing accuracy as revealed by small RNA deep sequencing analysis.Citation37 5′ capping and 3′ adenylation of pri-miRNA may play important roles for miRNA processing. Disruption of either 5′ capping or 3′ adenylation in cdkf;1 or cdkd123 mutant, respectively, leads to the less accumulation of mature miRNAs.Citation40 In addition, two cap-binding proteins, CBP20/80 have been shown to participate in pri-miRNA processing.Citation41,Citation42
miRNA Stability Control
After generation, miRNAs are 2’-O-methylated at 3′ terminal ribose by an RNA methyltransferase named HUA ENHANCER 1 (HEN1).Citation43 HEN1 methylates 21–24 nt RNA duplexes with 2 nt overhangs but not single-stranded RNAs (ssRNAs) in vitro.Citation43,Citation44 Crystal structure analyses of HEN1 reveal that the coordinated action of multiple RNA binding domains and the methyltransferase domain measures the length of the RNA duplex and determines the substrate specificity.Citation45 In hen1 mutant, miRNAs and small interfering RNAs (siRNAs, another class of endogenous non-coding small RNAs represses gene expression) are reduced in abundance and become heterogeneous in size.Citation46 Furthermore, siRNAs compete with miRNAs for HEN1 mediated methylation since depletion of siRNAs in weak alleles of hen1 increases overall methylated miRNAs levels.Citation47 The heterogeneity is caused by trimming and tailing activities at 3′ ends of the unmethylated small RNAs, while the 5′ ends are not affected by hen1 mutation.Citation46 Uridine is preferentially used for the addition of untemplated nucleotides and thus the tailing process is sometimes referred to as uridylation.Citation46 These results demonstrate that methylation protects miRNAs and siRNAs from 3′ degradation and uridylation.Citation46 Recently, both forward and reverse genetics identified HEN1 SUPRESSOR1 (HESO1) as an enzyme uridylating miRNAs in hen1.Citation48,Citation49 The facts that miRNA abundance is increased in heso1 hen1 and decreased in hen1 overexpressing HESO1 demonstrate that uridylation promotes degradation. The degradation should be highly processive as no degradation intermediates is detected in vivo. It’s worth noting that additional enzyme(s) responsible for miRNA uridylation must exist since miRNA U-tail is not abolished in hen1 heso1. However, T-DNA mutants of all other nine putative terminal nucleotidyl transferases failed to rescue hen1–8 phenotype, indicating that they may only play a minor role and their function in miRNA uridylation may be only detectable in the absence of HESO1. One effective way to test this is to introduce one or more of these mutations into hen1heso1. Also, a role of HESO1 in HEN1 competent background is unknown. It is possible that HESO1 acts on some small RNAs to regulate their stability and/or function. In addition, HESO1 may have RNA substrates other than small RNAs. In support of these, MUT68, also encodes a terminal nucleotidyl transferase, has dual roles in adenylating 5′ fragment of target RNA generated by AGO cleavage and uridylating unmethylated miRNAs in green algae Chlamydomonas reinhardtii.Citation50,Citation51
In hen1, the 3′-to-5′ truncated miRNAs are also observed. Their abundance is increased in hen1 heso1 relative to hen1. This result suggests that the 3′-to-5′ trimming activity most likely competes with U-tailing activity for unmethylated small RNAs. The enzymes responsible for 3′-to-5′ trimming in hen1 are currently unknown. A potential candidate is the small RNA-degrading nuclease (SDN), which degrades small RNAs from 3′-to-5′ in vitro.Citation52 Arabidopsis encodes a total of four SDN family proteins. Simultaneously knock-down of several SDN genes causes elevated miRNA levels and pleiotropic phenotypes, demonstrating that SDN has a role in miRNA turnover.Citation52 Another potential candidate for 3′-to-5′ trimming of miRNAs is homolog protein of Nibbler. Nibbler catalyzes 3′-to-5′ trimming of miRNAs in animals.Citation53,Citation54
Future Perspective
The core question in pri-miRNA processing is how DCL1 recognizes and processes pri-miRNAs precisely and efficiently. Analyses of the detailed function of DCL1 complex components will help us address this. A characteristic feature of DCL1 complex components is their ability of RNA binding. High-throughput sequencing of RNAs isolated by cross-linking immunoprecipitation (HITS-CLIP)Citation55 is a powerful tool for the whole genome identification of binding sites by these proteins. Recent studies have also uncovered diverse miRNAs modifications and their roles in stability control. A key challenge in the identification of enzymes in miRNA decay is that multiple enzymes often function in parallel and/or redundantly. In addition, some of the decay enzymes may be essential for multiple RNA degradation and knockout mutants may be non-viable, which makes them difficult to be discovered. In conclusion, the knowledge on the regulation of miRNA abundance will undoubtedly increase our ability to manipulate RNA silence and gene regulation for both agronomic and therapeutic purposes.
Acknowledgments
We thank Meng Xie, Xiang Liu and Shuxin Zhang from University of Nebraska-Lincoln, NE, USA for critical reading of the manuscript. This work was supported by the National Science Foundation (MCB-1121193).
References
- Llave C, Xie Z, Kasschau KD, Carrington JC. Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 2002; 297:2053 - 6; 10.1126/science.1076311; PMID: 12242443
- Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP. MicroRNAs in plants. Genes Dev 2002; 16:1616 - 26; 10.1101/gad.1004402; PMID: 12101121
- Park W, Li J, Song R, Messing J, Chen X. CARPEL FACTORY, a Dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana.. Curr Biol 2002; 12:1484 - 95; 10.1016/S0960-9822(02)01017-5; PMID: 12225663
- Zhang B, Pan X, Cannon CH, Cobb GP, Anderson TA. Conservation and divergence of plant microRNA genes. Plant J 2006; 46:243 - 59; 10.1111/j.1365-313X.2006.02697.x; PMID: 16623887
- Rhoades MW, Reinhart BJ, Lim LP, Burge CB, Bartel B, Bartel DP. Prediction of plant microRNA targets. Cell 2002; 110:513 - 20; 10.1016/S0092-8674(02)00863-2; PMID: 12202040
- Khraiwesh B, Zhu JK, Zhu J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim Biophys Acta 2012; 1819:137-48.
- Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek SH, et al. MicroRNA genes are transcribed by RNA polymerase II. EMBO J 2004; 23:4051 - 60; 10.1038/sj.emboj.7600385; PMID: 15372072
- Xie Z, Allen E, Fahlgren N, Calamar A, Givan SA, Carrington JC. Expression of Arabidopsis MIRNA genes. Plant Physiol 2005; 138:2145 - 54; 10.1104/pp.105.062943; PMID: 16040653
- Chen X. Small RNAs and their roles in plant development. Annu Rev Cell Dev Biol 2009; 25:21 - 44; 10.1146/annurev.cellbio.042308.113417; PMID: 19575669
- Voinnet O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009; 136:669 - 87; 10.1016/j.cell.2009.01.046; PMID: 19239888
- Rajagopalan R, Vaucheret H, Trejo J, Bartel DP. A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes Dev 2006; 20:3407 - 25; 10.1101/gad.1476406; PMID: 17182867
- Baumberger N, Baulcombe DC. Arabidopsis ARGONAUTE1 is an RNA Slicer that selectively recruits microRNAs and short interfering RNAs. Proc Natl Acad Sci U S A 2005; 102:11928 - 33; 10.1073/pnas.0505461102; PMID: 16081530
- Qi Y, Denli AM, Hannon GJ. Biochemical specialization within Arabidopsis RNA silencing pathways. Mol Cell 2005; 19:421 - 8; 10.1016/j.molcel.2005.06.014; PMID: 16061187
- Eamens AL, Smith NA, Curtin SJ, Wang MB, Waterhouse PM. The Arabidopsis thaliana double-stranded RNA binding protein DRB1 directs guide strand selection from microRNA duplexes. RNA 2009; 15:2219 - 35; 10.1261/rna.1646909; PMID: 19861421
- Smith MR, Willmann MR, Wu G, Berardini TZ, Möller B, Weijers D, et al. Cyclophilin 40 is required for microRNA activity in Arabidopsis. Proc Natl Acad Sci U S A 2009; 106:5424 - 9; 10.1073/pnas.0812729106; PMID: 19289849
- Iki T, Yoshikawa M, Meshi T, Ishikawa M. Cyclophilin 40 facilitates HSP90-mediated RISC assembly in plants. EMBO J 2012; 31:267 - 78; 10.1038/emboj.2011.395; PMID: 22045333
- Earley KW, Poethig RS. Binding of the cyclophilin 40 ortholog SQUINT to Hsp90 protein is required for SQUINT function in Arabidopsis. J Biol Chem 2011; 286:38184 - 9; 10.1074/jbc.M111.290130; PMID: 21908611
- Lobbes D, Rallapalli G, Schmidt DD, Martin C, Clarke J. SERRATE: a new player on the plant microRNA scene. EMBO Rep 2006; 7:1052 - 8; 10.1038/sj.embor.7400806; PMID: 16977334
- Yang L, Liu Z, Lu F, Dong A, Huang H. SERRATE is a novel nuclear regulator in primary microRNA processing in Arabidopsis. Plant J 2006; 47:841 - 50; 10.1111/j.1365-313X.2006.02835.x; PMID: 16889646
- Han MH, Goud S, Song L, Fedoroff N. The Arabidopsis double-stranded RNA-binding protein HYL1 plays a role in microRNA-mediated gene regulation. Proc Natl Acad Sci U S A 2004; 101:1093 - 8; 10.1073/pnas.0307969100; PMID: 14722360
- Fang Y, Spector DL. Identification of nuclear dicing bodies containing proteins for microRNA biogenesis in living Arabidopsis plants. Curr Biol 2007; 17:818 - 23; 10.1016/j.cub.2007.04.005; PMID: 17442570
- Song L, Han MH, Lesicka J, Fedoroff N. Arabidopsis primary microRNA processing proteins HYL1 and DCL1 define a nuclear body distinct from the Cajal body. Proc Natl Acad Sci U S A 2007; 104:5437 - 42; 10.1073/pnas.0701061104; PMID: 17369351
- Fujioka Y, Utsumi M, Ohba Y, Watanabe Y. Location of a possible miRNA processing site in SmD3/SmB nuclear bodies in Arabidopsis. Plant Cell Physiol 2007; 48:1243 - 53; 10.1093/pcp/pcm099; PMID: 17675322
- Dong Z, Han MH, Fedoroff N. The RNA-binding proteins HYL1 and SE promote accurate in vitro processing of pri-miRNA by DCL1. Proc Natl Acad Sci U S A 2008; 105:9970 - 5; 10.1073/pnas.0803356105; PMID: 18632569
- Liu C, Axtell MJ, Fedoroff NV. The helicase and RNaseIIIa domains of Arabidopsis Dicer-Like1 modulate catalytic parameters during microRNA biogenesis. Plant Physiol 2012; 159:748 - 58; 10.1104/pp.112.193508; PMID: 22474216
- Werner S, Wollmann H, Schneeberger K, Weigel D. Structure determinants for accurate processing of miR172a in Arabidopsis thaliana.. Curr Biol 2010; 20:42 - 8; 10.1016/j.cub.2009.10.073; PMID: 20015654
- Mateos JL, Bologna NG, Chorostecki U, Palatnik JF. Identification of microRNA processing determinants by random mutagenesis of Arabidopsis MIR172a precursor. Curr Biol 2010; 20:49 - 54; 10.1016/j.cub.2009.10.072; PMID: 20005105
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol 2009; 10:126 - 39; 10.1038/nrm2632; PMID: 19165215
- Cuperus JT, Montgomery TA, Fahlgren N, Burke RT, Townsend T, Sullivan CM, et al. Identification of MIR390a precursor processing-defective mutants in Arabidopsis by direct genome sequencing. Proc Natl Acad Sci U S A 2010; 107:466 - 71; 10.1073/pnas.0913203107; PMID: 20018656
- Song L, Axtell MJ, Fedoroff NV. RNA secondary structural determinants of miRNA precursor processing in Arabidopsis. Curr Biol 2010; 20:37 - 41; 10.1016/j.cub.2009.10.076; PMID: 20015653
- Addo-Quaye C, Snyder JA, Park YB, Li YF, Sunkar R, Axtell MJ. Sliced microRNA targets and precise loop-first processing of MIR319 hairpins revealed by analysis of the Physcomitrella patens degradome. RNA 2009; 15:2112 - 21; 10.1261/rna.1774909; PMID: 19850910
- Bologna NG, Mateos JL, Bresso EG, Palatnik JF. A loop-to-base processing mechanism underlies the biogenesis of plant microRNAs miR319 and miR159. EMBO J 2009; 28:3646 - 56; 10.1038/emboj.2009.292; PMID: 19816405
- Zhang W, Gao S, Zhou X, Xia J, Chellappan P, Zhou X, et al. Multiple distinct small RNAs originate from the same microRNA precursors. Genome Biol 2010; 11:R81; 10.1186/gb-2010-11-8-r81; PMID: 20696037
- Laubinger S, Zeller G, Henz SR, Buechel S, Sachsenberg T, Wang JW, et al. Global effects of the small RNA biogenesis machinery on the Arabidopsis thaliana transcriptome. Proc Natl Acad Sci U S A 2010; 107:17466 - 73; 10.1073/pnas.1012891107; PMID: 20870966
- Xie Z, Kasschau KD, Carrington JC. Negative feedback regulation of Dicer-Like1 in Arabidopsis by microRNA-guided mRNA degradation. Curr Biol 2003; 13:784 - 9; 10.1016/S0960-9822(03)00281-1; PMID: 12725739
- Pouch-Pélissier MN, Pélissier T, Elmayan T, Vaucheret H, Boko D, Jantsch MF, et al. SINE RNA induces severe developmental defects in Arabidopsis thaliana and interacts with HYL1 (DRB1), a key member of the DCL1 complex. PLoS Genet 2008; 4:e1000096; 10.1371/journal.pgen.1000096; PMID: 18551175
- Ren G, Xie M, Dou Y, Zhang S, Zhang C, Yu B. Regulation of miRNA abundance by RNA binding protein TOUGH in Arabidopsis. Proc Natl Acad Sci U S A 2012; 109:12817 - 21; 10.1073/pnas.1204915109; PMID: 22802657
- Yu B, Bi L, Zheng B, Ji L, Chevalier D, Agarwal M, et al. The FHA domain proteins DAWDLE in Arabidopsis and SNIP1 in humans act in small RNA biogenesis. Proc Natl Acad Sci U S A 2008; 105:10073 - 8; 10.1073/pnas.0804218105; PMID: 18632581
- Calderon-Villalobos LI, Kuhnle C, Dohmann EM, Li H, Bevan M, Schwechheimer C. The evolutionarily conserved TOUGH protein is required for proper development of Arabidopsis thaliana. Plant Cell 2005; 17:2473 - 85; 10.1105/tpc.105.031302; PMID: 16024589
- Hajheidari M, Farrona S, Huettel B, Koncz Z, Koncz C. CDKF;1 and CDKD protein kinases regulate phosphorylation of serine residues in the C-terminal domain of Arabidopsis RNA polymerase II. Plant Cell 2012; 24:1626 - 42; 10.1105/tpc.112.096834; PMID: 22547781
- Kim S, Yang JY, Xu J, Jang IC, Prigge MJ, Chua NH. Two cap-binding proteins CBP20 and CBP80 are involved in processing primary MicroRNAs. Plant Cell Physiol 2008; 49:1634 - 44; 10.1093/pcp/pcn146; PMID: 18829588
- Laubinger S, Sachsenberg T, Zeller G, Busch W, Lohmann JU, Rätsch G, et al. Dual roles of the nuclear cap-binding complex and SERRATE in pre-mRNA splicing and microRNA processing in Arabidopsis thaliana. Proc Natl Acad Sci U S A 2008; 105:8795 - 800; 10.1073/pnas.0802493105; PMID: 18550839
- Yu B, Yang Z, Li J, Minakhina S, Yang M, Padgett RW, et al. Methylation as a crucial step in plant microRNA biogenesis. Science 2005; 307:932 - 5; 10.1126/science.1107130; PMID: 15705854
- Yang Z, Ebright YW, Yu B, Chen X. HEN1 recognizes 21-24 nt small RNA duplexes and deposits a methyl group onto the 2′ OH of the 3′ terminal nucleotide. Nucleic Acids Res 2006; 34:667 - 75; 10.1093/nar/gkj474; PMID: 16449203
- Huang Y, Ji L, Huang Q, Vassylyev DG, Chen X, Ma JB. Structural insights into mechanisms of the small RNA methyltransferase HEN1. Nature 2009; 461:823 - 7; 10.1038/nature08433; PMID: 19812675
- Li J, Yang Z, Yu B, Liu J, Chen X. Methylation protects miRNAs and siRNAs from a 3′-end uridylation activity in Arabidopsis. Curr Biol 2005; 15:1501 - 7; 10.1016/j.cub.2005.07.029; PMID: 16111943
- Yu B, Bi L, Zhai J, Agarwal M, Li S, Wu Q, et al. siRNAs compete with miRNAs for methylation by HEN1 in Arabidopsis. Nucleic Acids Res 2010; 38:5844 - 50; 10.1093/nar/gkq348; PMID: 20448024
- Zhao Y, Yu Y, Zhai J, Ramachandran V, Dinh TT, Meyers BC, et al. The Arabidopsis nucleotidyl transferase HESO1 uridylates unmethylated small RNAs to trigger their degradation. Curr Biol 2012; 22:689 - 94; 10.1016/j.cub.2012.02.051; PMID: 22464194
- Ren G, Chen X, Yu B. Uridylation of miRNAs by hen1 suppressor1 in Arabidopsis. Curr Biol 2012; 22:695 - 700; 10.1016/j.cub.2012.02.052; PMID: 22464191
- Ibrahim F, Rymarquis LA, Kim EJ, Becker J, Balassa E, Green PJ, et al. Uridylation of mature miRNAs and siRNAs by the MUT68 nucleotidyltransferase promotes their degradation in Chlamydomonas. Proc Natl Acad Sci U S A 2010; 107:3906 - 11; 10.1073/pnas.0912632107; PMID: 20142471
- Ibrahim F, Rohr J, Jeong WJ, Hesson J, Cerutti H. Untemplated oligoadenylation promotes degradation of RISC-cleaved transcripts. Science 2006; 314:1893; 10.1126/science.1135268; PMID: 17185594
- Ramachandran V, Chen X. Degradation of microRNAs by a family of exoribonucleases in Arabidopsis. Science 2008; 321:1490 - 2; 10.1126/science.1163728; PMID: 18787168
- Han BW, Hung JH, Weng Z, Zamore PD, Ameres SL. The 3′-to-5′ exoribonuclease Nibbler shapes the 3′ ends of microRNAs bound to Drosophila Argonaute1. Curr Biol 2011; 21:1878 - 87; 10.1016/j.cub.2011.09.034; PMID: 22055293
- Liu N, Abe M, Sabin LR, Hendriks GJ, Naqvi AS, Yu Z, et al. The exoribonuclease Nibbler controls 3′ end processing of microRNAs in Drosophila. Curr Biol 2011; 21:1888 - 93; 10.1016/j.cub.2011.10.006; PMID: 22055292
- Darnell RB. HITS-CLIP: panoramic views of protein-RNA regulation in living cells. Wiley Interdiscip Rev RNA 2010; 1:266 - 86; 10.1002/wrna.31; PMID: 21935890