
Abstract
Reactive oxygen species (ROS) are involved in various cellular processes in plants. Among those, resistance to abiotic stress, defence mechanisms and cell expansion have been intensively studied during the last years. We recently demonstrated that ROS, in concert with auxin, have a role in cell cycle activation of differentiated leaf cells.1 In this addendum we provide further evidence to show that oxidative stress/ROS accelerate auxin-mediated cell cycle entry (G0-to-G1
Addendum to: Pasternak TP, Ötvös K, Domoki M, Fehér A. Linked activation of cell division and oxidative stress defense in alfalfa leaf protoplast-derived cells is dependent on exogenous auxin. Plant Growth Regul 2007; 51:109-17.
Figures and Tables
Figure 1 Oxidative stress/ROS enhance and are required for G0-to-G1 transition in leaf protoplast-derived alfalfa cells as indicated by nuclear parameters. (A) Diphenylene iodonium (DPI; 1 µM) applied to leaf protoplasts prevents cell activation in the presence of auxin (1 µM 2,4-dichlorophenoxyacetic acid, 2,4-D) and cytokinin (1 µM zeatin) as indicated by flow cytometric analysis (Part a) and microscopic investigation (Part b) of PI-stained (4 µg/ml; staining DNA as well as RNA) nuclei at four days of culture. (Part a) The histograms show the numbers of nuclei falling into fluorescence intensity groups (as defined by detection “channels”) depending on the amount and accessibility of nuclear DNA and RNA for staining. The fluorescence channels representing nuclei in G0 and G1 phases are indicated. (Part b) The sizes of nuclei and nucleoli in DPI-treated and control cells, as above. Increased sizes indicate active transcription and rRNA synthesis. DPI-treatment kept the nuclei as they could be observed in leaf cells (data not shown). DPI-DPI + auxin, A-auxin. (B) Copper stress (50 µM CuSO4) enhanced cell activation in the presence of 1 µM 2,4-D (and 1 µM zeatin). Alfalfa leaf protoplasts were cultured without auxin and copper (P), with copper stress (Cu), or copper stress and auxin (A + Cu). Nuclei were subjected to flow cytometric analysis following propidium iodide (PI, DNA and RNA) staining without (Part a) or after RNase-treatment (Part b) as above. RNase treatment diminished the difference of nuclear fluorescence among the control and treated cells indicating the role of nuclear RNA accumulation in the fluorescence intensity differences. Microscopic observations (Part c) of stained (DAPI staining DNA, left; PI staining DNA and RNA, right) highlighted the differences among cells treated by copper in the presence or absence of auxin, especially in the size and RNA content (arrow) of the nucleoli and homogeneity of nucleoplasm staining.
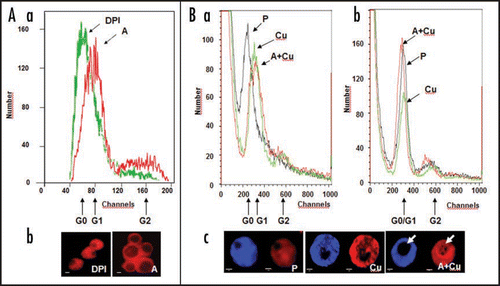
Figure 2 Effect of copper stress and DPI application on the cell cycle re-entry of stationary phase cultured plant cells. (A) The amount and activity of the cell cycle-regulatory CDKA/BMs kinase in 11-day old suspension-cultured alfalfa cells following subculture into fresh medium supplemented with copper (30 µM CuSO4, Cu), DPI (1 µM, DPI), their combination (Cu + DPI), or nothing (Contr.). The upper histogram shows the densitometry values of the CDK activity (as in vitro histone H1 phosphorylation activity of immunoprecipitated CDKA/BMs protein complexes also shown in the upper lane as an autoradiogram). The CDKA/BMs protein amount is determined by Western analysis (lower lane). (B) Growth of the control and treated cells as shown by fresh weight four days after subculture. The initial cell density is shown as a dashed line.
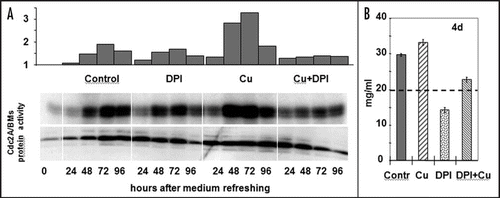
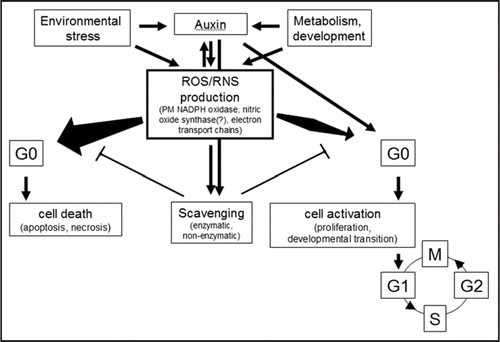
Figure 3 The scheme of potential interactions determining cellular response to oxidative stress. ROS/RNS are formed in the cells as the part of normal metabolism as well as in response to environmental stresses. ROS/RNS themselves as well as growth regulators regulate scavenging pathways to keep ROS/RNS levels low (for simplicity, only auxin is shown as beeing the main growth regulator implicated in cell cycle activation). In case net ROS/RNS production exceeds a limit due to increased production or decreased removal, differentiated cells will die. In contrast, a transient and moderate increase in ROS/RNS level may enhance cell activation in concert with auxin action.
Addendum to: