Abstract
Glutamate is of central importance in plant N metabolism since the biosynthesis of all other amino acids requires this compound. Glutamate dehydrogenase (GDH; EC 1.4.1.2), which catalyzes in vitro reversible reductive amination of 2-oxoglutatre to form glutamate, is a key player in the metabolism of glutamate. While most previous studies have indicated that the oxidative deamination is the in vivo direction of the GDH reaction, its physiological role has remained ambiguous for decades. We have recently isolated mutants for the two known Arabidopsis GDH genes and created a gdh double mutant. Our recent work revealed an increased susceptibility of the gdh double mutant to dark-induced C starvation, the first phenotype associated with the loss of GDH activity in plants. Monitoring the amino acid breakdown during the dark treatment also suggested that the deamination of glutamate catalyzed by GDH is central to the catabolism of many other amino acids.
Addendum to: Miyashita Y, Good AG. NAD(H)-dependent glutamate dehydrogenase is essential for the survival of Arabidopsis thaliana during dark-induced carbon starvation. J Exp Bot 2008; 59:667-80.
Nitrogen is a major limiting factor for plant growth and development and therefore the incorporation of inorganic N into amino acids is of central importance in agriculture. GDH had been viewed as the major ammonium assimilatory enzyme in the past. However, the discovery of glutamine synthetase (GS)/glutamate synthase (GOGAT) pathway as the sole port of ammonium assimilation raised the long-lasting question regarding the role played by GDH.Citation1,Citation2 This was confounded by the uncertainty about the in vivo direction of the GDH reaction, which reversibly aminates 2-oxoglutatrate or deaminates glutamate in vitro. Plants contain at least two GDH genes encoding either the α or β subunit of GDH.Citation3 Random assembling of the two subunits in the hexameric holoenzyme results in the seven isoenzymes which can be visualized on a non-denaturing gel [β6 (isoenzyme 1), α1β5 … α5β1, α6 (isoenzyme 7)].Citation4 For the in vivo direction that GDH catalyzes, oxidative deamination has been supported by many labeling experiments.Citation2 In two recent studies, the overexpression of either α or β subunit of GDH resulted in the increased release of 15NH4+ after [15N]glutamate feeding, demonstrating the deaminating role of the distinct GDH isoenzymes.Citation5,Citation6 In contrast, 15NH4+ incorporation into glutamate through GDH was demonstrated under salt stress in which continued 15NH4+ assimilation occurred in the presence of the GS inhibitor, methionine sulfoximine.Citation7 Nevertheless, most previous results supported the current consensus that GDH deaminates glutamate in vivo under normal growth conditions.
Although previous labeling experiments were useful to define the in vivo direction of the GDH reaction, the physiological role of GDH remained speculative. For this reason, GDH expression has been extensively studied at the transcript, protein and enzyme activity levels. In these previous studies, the regulation of GDH by light/dark cycle, sugar levels and ammonia addition was documented.Citation2 In brief, expression of GDH appears to be controlled by the cellular energy state or sugar levels since the dark-induction of GDH can be repressed by exogenous addition of sucrose.Citation3,Citation8 This tight regulation of GDH according to the cellular C status has led researchers to suggest that GDH is involved in supplying an alternative C source to the respiratory pathway during sugar starvation.Citation2,Citation8 However, this hypothesis has not been demonstrated until recently, since the previously characterized gdh mutants as well as the transgenic plants overexpressing GDH did not exhibit a visible phenotype that supports the proposed role of GDH.Citation5,Citation6,Citation8–Citation10 In particular, all previous gdh mutants contained mutations only in one of the two GDH genes.Citation8–Citation10 It was possible that the effects of the loss of one functional GDH were masked by the redundant function of the other GDH in previous studies. Recently, we have isolated mutants for both of the two known Arabidopsis GDH genes and obtained a gdh double mutant (gdh1–2/gdh2-1) from crossing gdh1 and gdh2 mutants. Using these mutants, we tested the role of GDH during sugar starvation by subjecting the plants to prolonged darkness. In this experiment, we observed an increased susceptibility of gdh1–2/gdh2-1 to prolonged darkness. The recovery of the phenotype of gdh1–2/gdh2-1 by the addition of sucrose before the onset of the dark treatment indicated that the dark-induced C starvation was the cause of the phenotype. In addition, absence of the phenotype in both of the single mutants suggested the functional redundancy of the two GDH genes. Based on the view that GDH deaminates glutamate in vivo, the anaplerotic GDH reaction functions to funnel the C skeleton of glutamate into the TCA cycle during sugar starvation.
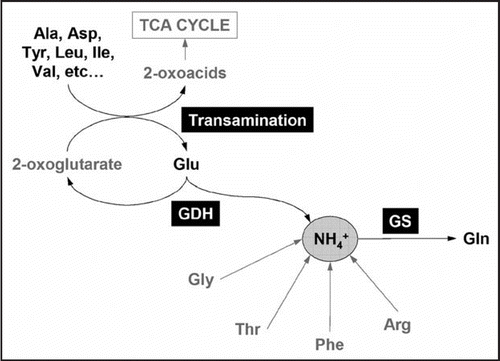
Quantification of amino acid levels also revealed the profound effects of the loss of GDH activity on amino acid metabolism. In general, sugar starvation accelerates the protein degradation through the autophagic process, resulting in the increase in free amino acids.Citation11 Under such conditions, catabolism of the accumulated free amino acids becomes important as the source of respiratory substrates. In our study, levels of several amino acids significantly differed between WT and the gdh mutants during the dark treatment although the amino acid profile of WT and those of the gdh mutants were similar under normal conditions. During the dark treatment, levels of alanine and aspartate decreased in the controls, but conversely, increased in the gdh mutants. Also, an increased accumulation of branched- chain amino acids (i.e., leucine, isoleuccine and valine) was observed in gdh1–2/gdh2-1. Interestingly, increase and decrease in glutamate levels were observed during the dark treatment in controls and gdh1–2/gdh2-1, respectively. Based on these observations, we proposed that glutamate deamination by GDH is essential for the generation of 2-oxoglutate, which acts as an amino group acceptor in the transaminations required for the catabolism of several amino acids (). The amino acid species over-accumulated in gdh1–2/gdh2-1 during the dark treatment were in fact those that can be converted to respirable 2-oxoacids through the transaminations. Co-localization of GDH and various aminotransferases in the mitochondriaCitation12 further supports the proposed model. Since the deamination of amino acids can be catalyzed by only a handful of enzymes in plants (), glutamate deamination by GDH may be central in amino acid catabolism.
The proposed role of GDH in amino acid catabolism also indicates the potential involvement of GDH in N remobilization during senescence. In fact, induction of GDH in senescing leaves has been documented in plants.Citation13,Citation14 Since the dark-induced C starvation mimics the physiological process of senescence, deamination catalyzed by GDH appears to be important source of ammonia that can be re-assimilated into N transport compounds (glutamine and asparagine). This potential role of GDH during senescence is particularly interesting since cytosolic GS1, which is induced during senescence, has been shown to be a major component that determines the grain production in maize.Citation15 The co-expression of cytosolic GS1 and GDH during senescenceCitation13,Citation14 points to an efficient pathway for the synthesis of N transport compounds from the free amino acids derived from proteolysis ().
Our recent work gave insights into the physiological role of GDH in amino acid catabolism in plants. However, further investigations are necessary to more precisely define the role of this long-disputed enzyme.
Figures and Tables
Figure 1 Proposed model of amino acid catabolism during sugar starvation. Deamination of glutamate is catalyzed by GDH. The enzymes such as glycine decarboxylase, threonine deaminase, phenylalanine lyase and arginase also catalyze the deamination of the corresponding amino acids. Among these deaminating reactions, GDH appears to be the major route for amino acid breakdown as the amino group of other amino acids can be transferred to Glu through transaminations. The released ammonia can be re-assimilated into Gln by glutamine synthetase (GS). During senescence, Gln can be translocated to sink tissues.
Addendum to:
References
- Miflin BJ, Habash D. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J Exp Bot 2002; 53:979 - 987
- Dubois F, Tercé Laforgue T, Gonzalez Moro MB, Estavillo MB, Sangwan R, Gallais A, Hirel B. Glutamate dehydrogenase in plants: is there a new story for an old enzyme?. Plant Physiol Biochem 2003; 41:565 - 576
- Purnel MP, Skopelitis DS, Roubelakis Angelakis KA, Botella JR. Modulation of higher-plant NAD(H)-dependent glutamate dehydrogenase activity in transgenic tobacco via alteration of beta subunit levels. Planta 2005; 222:167 - 180
- Loulakakis KA, Roubelakis Angelakis KA. Plant NAD(H) glutamate dehydrogenase consists of two subunit polypeptides and their participation in the seven isoenzymes occurs in an ordered ratio. Plant Physiol 1991; 97:104 - 111
- Purnel MP, Botella JR. Tobacco isoenzyme 1 of NAD(H)-dependent glutamate dehydrogenase catabolizes glutamate in vivo. Plant Physiol 2007; 143:530 - 539
- Skopelitis DS, Paranychianakis NV, Kouvarakis A, Spyros A, Stephanou EG, Roubelakis Angelakis KA. The isoenzyme 7 of tobacco NAD(H)-dependent glutamate dehydrogenase exhibits high deaminating and low aminating activities in vivo. Plant Physiol 2007; 145:1726 - 1734
- Skopelitis DS, Paranychianakis NV, Paschalidis KA, Pliakonis ED, Yakoumakis D, Delis ID, Kouvarakis A, Papadakis AK, Stephanou E, Roubelakis-Angelakis KA. Abiotic stress generated ROS signal expression of anionic glutamate dehydrogenase to form glutamate for proline synthesis. Plant Cell 2006; 18:2767 - 2781
- Melo Oliveira R, Oliveira IC, Coruzzi GM. Arabidopsis mutant analysis and gene regulation define a nonredundant role for glutamate dehydrogenase in nitrogen assimilation. Proc Natl Acad Sci USA 1996; 93:4718 - 4723
- Magalhaes JR, Ju GC, Rich PJ, Rhodes D. Kinetics of 15NH4+ assimilation in Zea maize: preliminary studies with a glutamate dehydrogenase (GDH1) null mutant. Plant Physiol 1990; 94:647 - 656
- Fontaine JX, Saladino F, Agrimonti C, Bedu M, Tercé Laforgue T, Tetu T, Hirel B, Restivo RM, Dubois F. Control of the synthesis and subcellular targeting of the two GDH gene products in leaves and stems of Nicotiana plumbaginifolia and Arabidopsis thaliana. Plant Cell Physiol 2006; 47:410 - 418
- Yu SM. Cellular and genetic responses of plants to sugar starvation. Plant Physiol 1999; 121:687 - 693
- Liepman AH, Olsen LJ. Genomic analysis of aminotransferases in Arabidopsis thaliana. Crit Rev Plant Sci 2004; 23:73 - 89
- Masclaux C, Valadier MH, Brugière N, Morot Gaudry JF, Hirel B. Characterization of the sink/source transition in tobacco (Nicotiana tabacum L.) shootsin relation to nitrogen management and leaf senescence. Planta 2000; 211:510 - 518
- Masclaux Daubresse C, Reisdorf Cren M, Pageau K, Lelandais M, Grandjean O, Kronenberger J, Valadier MH, Feraud M, Jouglet T, Suzuki A. Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct role in the sink-source nitrogen cycle in tobacco. Plant Physiol 2006; 140:444 - 456
- Martin A, Lee J, Kichey T, Gerentes D, Zivy M, Tatout C, Dubois F, Balliau T, Valot B, Davanture M, Tercé Laforgue T, Quilleré I, Coque M, Gallais A, Gonzalez Moro MB, Bethencourt L, Habash DZ, Lea PJ, Charcosset A, Perez P, Murigneux A, Sakakibara H, Edwards KJ, Hirel B. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. Plant Cell 2006; 18:3252 - 3274