
Abstract
Receptor-like proteins (RLPs) are cell surface receptors that play important roles in various processes. In several plant species RLPs have been found to play a role in disease resistance, including the tomato Cf and Ve proteins and the apple HcrVf proteins that mediate resistance against the fungal pathogens Cladosporium fulvum, Verticillium spp. and Venturia inaequalis, respectively. The Arabidopsis genome contains 57 AtRLP genes. Two of these, CLV2 (AtRLP10) and TMM (AtRLP17), have well-characterized functions in meristem and stomatal development, respectively, while AtRLP52 is required for defense against powdery mildew. We recently reported the assembly of a genome-wide collection of T-DNA insertion lines for the Arabidopsis AtRLP genes. This collection was functionally analyzed with respect to plant growth, development and sensitivity to various stress responses including pathogen susceptibility. Only few new phenotypes were discovered; while AtRLP41 was found to mediate abscisic acid sensitivity, AtRLP30 (and possibly AtRLP18) was found to be required for full non-host resistance to a bacterial pathogen. Possibly, identification of novel phenotypes is obscured by functional redundancy. Therefore, RNA interference (RNAi) to target the expression of multiple AtRLP genes simultaneously was employed followed by functional analysis of the RNAi lines.
Addendum to: Wang G, Ellendorff U, Kemp B, Mansfield JW, Forsyth A, Mitchell K, Bastas K, Liu CM, Woods-Tör A, Zipfel C, de Wit PJGM, Jones JDG, Tör M, Thomma BPHJ. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol 2008; 147:503-17.
Acknowledgements
B.P.H.J.T. is supported by a Vidi grant of the Research Council for Earth and Life sciences (ALW) of the Netherlands Organization for Scientific Research (NWO). This project was cofinanced by the Centre for BioSystems Genomics (CBSG) which is part of the Netherlands Genomics Initiative/Netherlands Organisation for Scientific Research.
Figures and Tables
Figure 1 Cloning strategy for RNAi constructs. (A) PCR fragments of specific AtRLP fragments are generated with 5′HindIII (or BamHI for RNAi construct 2 and 5) and 3′ NotI and EcoRI restriction sites. (B) Inverted repeat constructs are generated by ligating HindIII (or BamHI for RNAi constructs 2 and 5) and NotI digested PCR fragment and HindIII (or BamHI for RNAi constructs 2 and 5) and EcoRI digested PCR fragment together with a NotI- and EcoRI-digested 129 bp spacer segment from the Pichia pastoris Aox-1 gene into the HindIII-digested (or BamHI for RNAi constructs 2 and 5) pGreen backbone. The fragments are not drawn to scale.
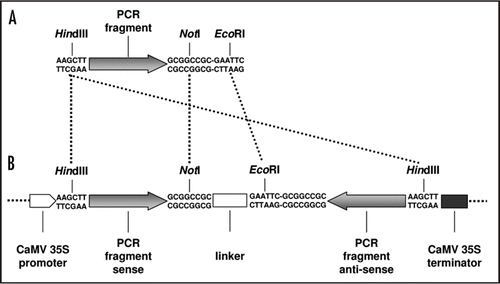
Figure 2 RNAi construct 5 triggers ABA-induced chlorosis. Comparison of the leaf phenotype of six independent transgenic lines containing RNAi construct 5 (a to f) with the parental line Col-0 three days after application of 100 mM abscisic acid (ABA).

Table 1 RNAi constructs to target homologous AtRLP genes
Addendum to: