Abstract
Phosphoinositides (PIs) control various cellular functions of eukaryotic cells. PIs are derived from phosphatidylinositol (PtdIns) by phosphorylation of the inositol-ring in the lipid-head group; the action of specific lipid kinases gives rise to a family of structurally-related PIs representing PtdIns-mono-, bis-, and -trisphosphates. Specific PIs, such as phosphatidylinositol-4,5-bisphosphate (PtdIns(4,5)P2), can influence more than one physiological process, raising the question as to how interactions with alternative protein partners are coordinated. Previous studies have proposed that PIs are organized by spatiotemporal compartmentation into distinct functional pools, however, mechanisms for the generation and maintenance of such pools has not been presented. Several recent studies now indicate that not only the distinctive inositolpolyphosphate head groups may be relevant for PI function but also the associated fatty acyl-moieties, which may be involved in sorting of lipid precursors into distinct pools. This mini-review aims at highlighting recent evidence that PI acyl-groups exert relevant effects on signaling.
Acknowledgements
The author would like to thank Prof. Dr. Ivo Feussner for helpful discussion and acknowledges generous financial support through an Emmy Noether grant from the German Research Foundation (DFG) and the Göttingen Graduate School for Neurosciences and Molecular Biosciences (GGNB).
Figures and Tables
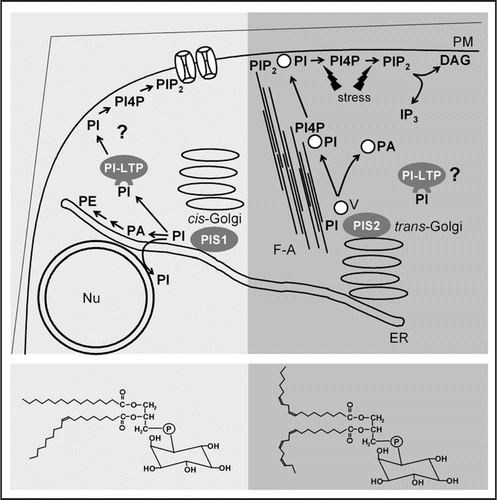
Figure 1 A model of functional phosphoinositide-pools in plant cells. All phosphoinositides originate from phosphatidylinositol (PI) formed by PI-synthases (PIS). The Arabidopsis isoenzymes, PIS1 and PIS2, differ in substrate specificity and localization, giving rise to PI-species with more saturated (bottom left) or more unsaturated fatty acids (bottom right). Localization of PIS1 and PIS2 with cis-Golgi and trans-Golgi sites is hypothetical. PI may be delivered to subcellular destinations by vesicle trafficking or by lipid transfer proteins. Left, Based on lipid patterns observed in Arabidopsis plants overexpressing PIS1,Citation23 PI formed by PIS1 may not enter the secretory pathway and instead support ER-associated PI-pools that may also be continuous with the nuclear envelope. In the absence of sufficient lipid transfer capacity, PI formed by overexpressed PIS1 may accumulate in the ER, followed by degradation to PA and increased PE production.Citation23 A physiological destination of PI-LTPs may be the plasma membrane, where intact saturated PI may be involved in the regulation of ion channels. Right, As PI-kinases and PIPkinases act on secretory vesicles and PIS2 overexpression resulted in increased formation of polyphosphoinositides,Citation23 PI formed by PIS2 may associate with trans-Golgi vesicles and experience partial conversion to PI4P and PIP2. PI delivered to the plasma membrane by vesicle-flow along F-actin strands may be converted by stress-activated PI-kinases and PIP-kinases; PI4P and PIP2 formed may serve as substrates for PLC and support IP3-production.Citation15 PI-LTPs may play a role in the delivery of unsaturated PI to subcellular destinations. For better clarity, short-hand abbreviations for the lipids were used for the model. ER, endoplasmic reticulum; F-A, filamentous actin; Nu, nucleus; PA, phosphatidic acid; PE, phosphatidylethanolamine; PI, phosphatidylinositol; PI4P, phosphatidylinositol-4-phosphate; PI-LTP, phosphoinositide-specific lipid transfer protein; PIP2, phosphatidylinositol-4,5-bisphosphate; PLC, phospholipase C; PM, plasma membrane; V, vesicle.