Abstract
The conserved eukaryotic protein SGT1 (Suppressor of G2 allele of skp1) participates in diverse physiological processes such as cell cycle progression in yeast, plant immunity against pathogens and plant hormone signalling. Recent genetic and biochemical studies suggest that SGT1 functions as a novel co-chaperone for cytosolic/nuclear HSP90 and HSP70 molecular chaperones in the folding and maturation of substrate proteins. Since proteins containing the leucine-rich repeat (LRR) protein-protein interaction motif are overrepresented in SGT1-dependent phenomena, we consider whether LRR-containing proteins are preferential substrates of an SGT1/HSP70/HSP90 complex. Such a chaperone organisation is reminiscent of the HOP/HSP70/HSP90 machinery which controls maturation and activation of glucocorticoid receptors in animals. Drawing on this parallel, we discuss the possible contribution of an SGT1-chaperone complex in the folding and maturation of LRR-containing proteins and its evolutionary consequences for the emergence of novel LRR interaction surfaces.
Figures and Tables
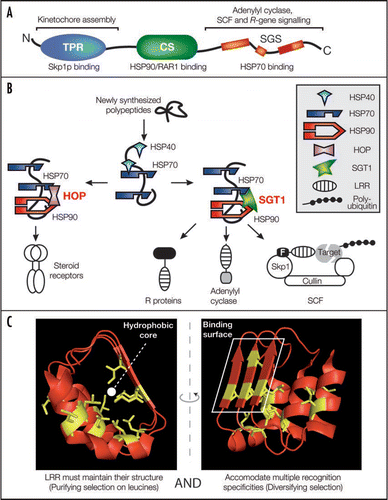
Figure 1 Model for SGT1/chaperone complex functions in the folding of LRR-containing proteins. (A) The structural domains of SGT1, their sites of action (above) and respective binding partners (below) are shown. N- and C-termini are indicated. TPR, tetratricopeptide repeat; CS, CHORD/SGT1; SGS, SGT1-specific. (B) Conceptual analogy between steroid receptor folding by the HOP/chaperone machinery and LRR protein folding by the SGT1/chaperone machinery. LRR motifs are overrepresented in processes requiring SGT1 such as plant immune receptor signalling, yeast adenylyl cyclase activity and plant or yeast SCF (Skp1/Cullin/F-box) E3 ubiquitin ligase activities. (C) Opposite forces drive LRR evolution. Structure of LRRs 16 to 18 of the F-box auxin receptor TIR1 is displayed as an illustration of the LRR folds.Citation30 Leucine/isoleucine residues (side chain displayed in yellow) are under strong purifying selection and build the hydrophobic LRR backbone (Left). By contrast, solvent-exposed residues of the β-strands define a polymorphic and hydrophilic binding surface conferring substrate specificity to the LRR (Right) and are often under diversifying selection.