
Abstract
Transmission of plant viruses is the result of interactions between a given virus, the host plant, and the vector. Most research has focused on molecular and cellular virus-vector interactions, and the host has only been regarded as a reservoir from which the virus is acquired by the vector more or less accidentally. However, a growing body of evidence suggests that the host can play a crucial role in transmission. Indeed, at least one virus, Cauliflower mosaic virus, exploits the host's cellular pathways to form specialized intracellular structures that optimize virus uptake by the vector and hence transmission.
Figures and Tables
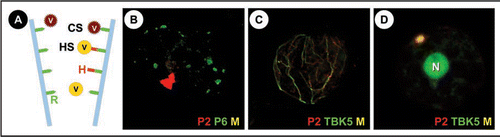
Figure 1 (A) The different strategies of non-circulant transmission: Viruses (V) using the capsid strategy (CS) attach directly to a receptor (R) in the tip of the a proboscis forming aphid stylets (blue), whereas in the helper strategy (HS) this interaction is mediated by the viral helper protein (H) that binds the virus particle to the receptor. Note that the helper protein can bind independently of the virus to the stylets. Whether the same receptor is used by different viruses as presented in the schema, is not known. (B) A turnip protoplast transfected with CaMV was double-labelled late in infection for CaMV helper protein P2 (red) and the marker protein for the virus factories P6 (green). It is visible that P2 localizes in a single, large transmission body, whereas the numerous virus factories are devoid of P2 (Colocalization would be revealed by yellowish color (M) in this superposition). (C and D) Turnip protoplasts were cotransfected with CaMV and TBK5-GFP and immunolabelled for P2 (red) and TBK5 was detected by GFP fluorescence. (C) shows a cell early in infection, where P2 and TBK5-GFP colocalize on a network that we identified as the microtubule cytoskeleton (unpublished data). (D) shows a cell later in infection where P2 and TBK5-GFP colocalize, as indicated by the yellowish color, in a transmission body. Note that TBK5-GFP also strongly labels the nucleus (N).
Addendum to: