
Abstract
Lateral root (LR) stimulation during early signal exchange between plant roots and ectomycorrhizal (ECM) fungi has recently been shown to be achieved by modulation of auxin gradients. We suggested that this modulation could occur through altered polar auxin transport (PAT) and through activation of auxin signalling pathways in the root. However, it remains unclear, which fungal molecules alter auxin pathways inside the plant partner. It has been suggested in previous studies that auxin released by the fungus could trigger observed plant responses during early signal exchange and later on during root colonization. Here we focus on the early interaction and we provide evidence for an alternative mechanism. Indeed, LR stimulation by the fungus in A. thaliana followed a totally different timing than with exogenously applied auxin. Furthermore, experimental conditions that excluded the exchange of soluble molecules while allowing exchange of volatile(s) between the plant and the fungus were sufficient for LR induction, therefore questioning the role of secreted fungal auxin. These data suggest that volatiles released by the fungus and sensed by the plant may act upstream of altered auxin signalling in the plant.
Acknowledgements
We are grateful to Krista Plett for proofreading the final version of the manuscript. We thank Zachary Gaudin for help with experiments and Francis Martin for critical reading. This work has been supported by the European Commission within the Seventh Framework Program for Research, Project Energypoplar (FP7-211917), the French Space Agency (CNES) and the EVOLTREE network of excellence.
Figures and Tables
Figure 1 Comparison of LR stimulation in Arabidopsis thaliana in contact with Laccaria bicolor and during exogenous IAA treatment. (A) LR development on 5 DAG Arabidopsis seedlings covered by a cellophane membrane with or without fungal mycelium or in the presence of 1 or 10 µM IAA. IAA rapidly stimulated LR development but led after two days to a plateau whereas LR stimulation by Laccaria is slower but persists over the first ten days. (B) LR development after 8 days of 10 µM IAA treatment. (C) LR development after 8 days indirect contact in the presence of 10 µM IAA. Note the high number of second degree LRs in (C) (arrows) that are absent in (B). LR stimulation in Arabidopsis by volatile molecules released by Laccaria mycelia. (D) Two-compartmented plate with Arabidopsis seedlings (left) and Laccaria (right). (E) LR development in the presence of volatiles released by Laccaria. Compared to controls, LR development was stimulated from three days of co-culturing with mycelium. Per treatment 15 to 25 (A–C) and 50 seedlings (E) were analyzed respectively. Different letters indicate significant difference between the respective conditions at each time-point (A and E) (Student's t-test, p < 0.05).

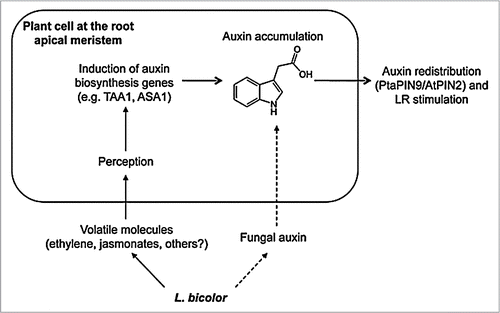
Figure 2 Hypothetical model of early L. bicolor/root signaling. The fungus releases some volatiles in addition to small amounts of auxin. Volatiles such as ethylene and jasmonate may be perceived by plant cells therefore activating the transcription of auxin biosynthesis genes, for instance Anthranilate synthase ASA1 or Tryptophane Amino Transferase TAA1. The activity of these genes could increase endogenous auxin levels in root apex. If present, fungal auxin may enrich this pool additionally. Increasing auxin levels may activate polar auxin transport and control redistribution of auxin, which will lead ultimately to LR stimulation.
Addendum to: