Abstract
Eukaryotic organisms rely on intricate signaling networks to connect recognition of microbes with the activation of efficient defense reactions. Accumulating evidence indicates that phospholipids are more than mere structural components of biological membranes. Indeed, phospholipid-based signal transduction is widely used in plant cells to relay perception of extracellular signals. Upon perception of the invading microbe, several phospholipid hydrolyzing enzymes are activated that contribute to the establishment of an appropriate defense response. Activation of phospholipases is at the origin of the production of important defense signaling molecules, such as oxylipins and jasmonates, as well as the potent second messenger phosphatidic acid (PA), which has been shown to modulate the activity of a variety of proteins involved in defense signaling. Here, we provide an overview of recent reports describing the different plant phospholipase pathways that are activated during the establishment of plant defense reactions in response to pathogen attack.
Figures and Tables
Figure 1 Schematic representation of phospholipid structure and the sites of cleavage by phospholipases. (A) General structure of a phospholipid, with two fatty acyl chains linked to a glycerol backbone, at the sn-1 and sn-2 positions, and a phosphate group creating the «phosphatidyl» moiety to which a variable head group is attached. The sites of phospholipase activity are indicated by arrows. Lipid acyl hydrolases (LAHs) are able to hydrolyze phospholipids at sn-1 and sn-2 positions. (B) Possible head groups and resulting phospholipids are indicated: PLA, phospholipase A; PLC, phospholipase C; PLD, phospholipase D; Fatty Acid 1, fatty acid in sn-1 position; Fatty Acid 2, fatty acid in sn-2 position; P, phosphate group.
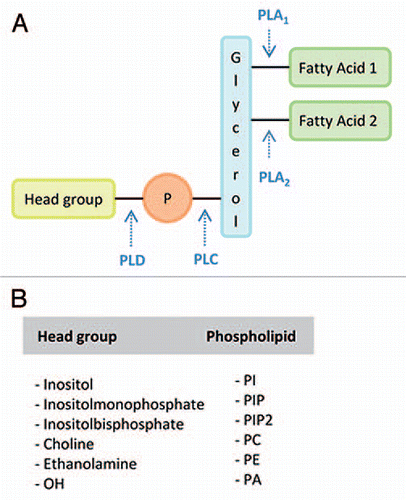
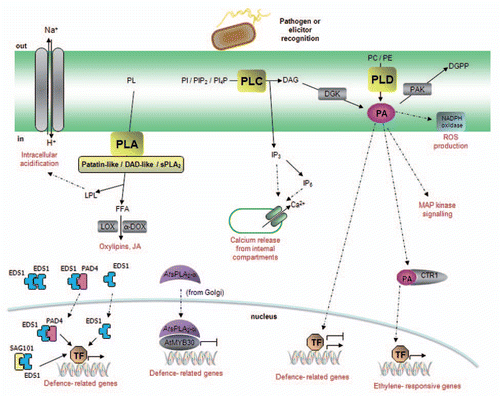
Figure 2 Model of phospholipase signaling in plant defense responses. Pathogen recognition induces the activation of phospholipase-mediated signaling pathways. Phospholipase A (PLA) hydrolyzes membrane phospholipids (PL) to produce free fatty acids (FFA) and lysophospholipids (LPL). PLAs are classified in three main groups: (1) the patatin-like proteins, responsible for the production of oxilipins and jasmonates, (2) the DAD (Defective in Anther Dehiscence)-like proteins with, to date, no well-established role in plant immunity and (3) the secretory PLA2s. Phospholipids from the plasma membrane, as well as from various intracellular membranes (mitochondria, Golgi, chloroplast…), are potential substrates for PLA activity. EDS1, PAD4 and SAG101 are essential immunity components that shuttle across the nuclear envelope to regulate defense responses. Despite their sequence homology to LAHs, the fact that no enzymatic activity has been demonstrated to date for these proteins suggests that their LAH domain may play a structural role rather than an enzymatic role. Similarly, a non-enzymatic function has been shown for AtsPLA2-α, a negative regulator of plant defense that is translocated into the nucleus in the presence of the transcription factor AtMYB30. Phospholipase C (PLC) is a plasma membrane protein that hydrolyzes phosphatidylinositol (PI), phosphatidylinositol bisphosphate (PIP2) or phosphatidylinositol phosphate (PI4P) to produce inositol trisphosphate (IP3) and diacylglycerol (DAG). IP3, which may be converted to IP6, diffuses in the cytosol where it releases Ca2+ from internal stores whereas DAG is rapidly converted to phosphatidic acid (PA) through the action of diacylglycerol kinase (DGK). Increased PA levels modulate additional signaling components, which are indicated in the figure. PA signaling may be attenuated via its phosphorylation to diacylglycerol pyrophosphate (DGPP) by a PA kinase (PAK). Finally, PLD also generates PA through the hydrolysis of structural phospholipids, such as phosphatidylcholine (PC) or phosphatidyl-ethanolamine (PE). PLDs present different subcellular localizations and may thus use phospholipids from the plasma membrane or from intracellular membranes.