Abstract
Light is the ultimate energy source for photo-autotrophs on earth. For green plants, however, it can also be toxic under certain stressful environmental conditions and at critical developmental stages. Anthocyanins, a class of flavonoids, act as an effective screening mechanism that allows plant survival and proliferation under occasional periods of harmful irradiation through modulation of light absorption. Apart from light-sensing through photoreceptors such as phytochrome and cryptochrome, plants use the photosynthetic electron transfer (PET) chain to integrate light information. The redox status of the plastoquinone (PQ) pool of the PET chain regulates anthocyanin biosynthesis genes, together with the plant hormone ethylene and plant hormone-like sugars. A complex signaling apparatus in acyanic cells appears to transduce information to cyanic cells to regulate anthocyanin production through an intercellular signaling pathway that remains largely uncharacterized. This review will highlight recent advances in this field and their implications for the regulation of anthocyanin pigmentation.
Acknowledgements
This work was supported by the Crop Functional Genomics Center (grant no. CG2151) and the NRF (grant no. 2009-008146) funded by the Ministry of Education, Science, and Technology, Korea, and by a Korea Research Foundation Fellowship (grant no. KRF-2006-C00083 to P.K.D.).
Figures and Tables
Figure 1 GUS expression in transgenic Arabidopsis seedlings harboring a GUS reporter gene driven by the PAP1 promoter (PAP1pro:GUS rep). Nine day-old seedlings were transferred to 0 (A) and 60 mM (B and C) Suc in the presence of 10 µM DCMU (C) and then exposed to white light (140 µmolm−2s−1) for 48 hours.
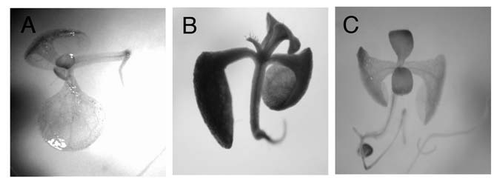
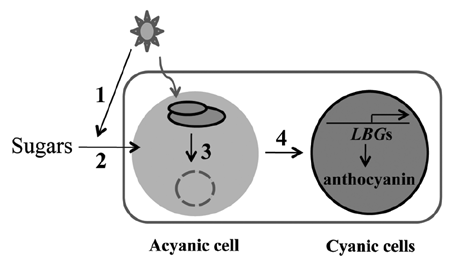
Figure 2 Proposed model for redox mediated signaling between cyanic and acyanic cells. (1) Light signals induce exogenous Suc uptake. (2) Apoplastic Suc in cotyledons and leaves is sensed by photosynthetic electron transport in mesophyll cells and generates a plastid signal. (3) The plastid signal is transformed into a mesophyll signal and then (4) transmitted to epidermal or vasculate cells, activating the MBW (Myb-bHLH-WD40) regulatory complex and downregulating MybL2 expression. This in turn leads to the specific upregulation of several late anthocyanin biosynthesis genes (LBGs), resulting in the accumulation of anthocyanin. The signaling molecule that mediates this intercellular response remains elusive.