Abstract
miRNAs, a subclass of small regulatory RNAs, are present from ancient unicellular protozoans to parasitic helminths and parasitic arthropods. The miRNA-silencing mechanism appears, however, to be absent in a number of protozoan parasites. Protozoan miRNAs and components of their silencing machinery possess features different from other eukaryotes, providing some clues on the evolution of the RNA-induced silencing machinery. miRNA functions possibly associate with neoblast biology, development, physiology, infection and immunity of parasites. Parasite infection can alter host miRNA expression that can favor both parasite clearance and infection. miRNA pathways are, thus, a potential target for the therapeutic control of parasitic diseases.
Introduction
microRNAs (miRNAs) are one subtype of small endogenous single-stranded RNAs and have been so far reported in many viruses, animals and plants such as Mareks diseased virus, fruit flies, humans, zebra fish and Arabidopsis. But the miRNA-induced silencing mechanism may be lost in yeast and some unicellular organisms.Citation1 miRNA-mediated regulation of gene expression may be pervasively distributed across the parasite kingdom, although miRNAs are absent in some unicellular parasites.Citation2,Citation3 Individual miRNAs can directly or indirectly repress hundreds or even thousands of genes, affecting protein production mostly on a fine scale via translational repression or/and mRNA degradation.Citation4,Citation5 A systematic survey showed that more than 20% human genes were regulated by miRNAs.Citation6 Although a study in which 95 miRNA genes were mutated in Caenorhabiditis elegans, no abnormal phenotypes were observed, indicating major miRNAs are not vital for the development or viability.Citation7 Increasing evidence supports the idea that miRNAs participate in complex regulatory networks and the abnormality of some miRNAs is related to occurrence of many human diseases, including cancer.Citation8-Citation12
Silencing of the main RNA-induced silencing complex (RISC) component was shown to promote the susceptibility of Anopheles gambiae to Plasmodium infection.Citation13 Similarly, dysfunction in Dicer and RNA-dependent RNA polymerase resulted in a loss of the regulation control of specific surface proteins in Giardia lamblia.Citation14 These observations indicate that regulatory small RNAs are essential in parasite infection. We herein outline our current understanding of miRNAs in unicellular and multicellular parasites and provide examples of some unique properties of protozoan miRNAs. The relationship between snoRNAs and miRNA precursors, miRNA markers for neoblasts and the potential roles of miRNAs in parasites are also discussed.
Biogenesis of miRNAs
In the canonical approach, miRNA genes are first transcribed into long pri-miRNA transcripts by RNA polymerase (Pol), usually Pol II. These pri-miRNAs are specifically spliced by Drosha, a nuclear RNase III, giving rise to the precursors of miRNAs with a size of approximately 70 nucleotides (pre-miRNAs). With the help of a nuclear transport receptor (exportin-5) that acts with the molecules with a stem and a short overhang at the 3′ end, the pre-miRNAs in the nucleus are transported to the cytoplasm. Afterwards, the molecules exported are cleaved into smaller duplexes, which are mediated by RNase III Dicer. To be functional, double-stranded shortened miRNAs, which are swiftly changed into single strand during assembly, are loaded onto an Argonaut-containing RNA-induced silencing complex (RISC).Citation15,Citation16 In most cases, the partner of the mature miRNA incorporated into the complex, named as miRNA*, is degraded. Recently, it has been shown that miRNAs* are also present at a relatively high level and have capacity of repressing targets ().Citation17 This finding adds the complexity of regulatory networks where miRNAs or miRNAs* dominate.
Figure 1. Biogenesis of canonical and mirtron miRNAs in animals.
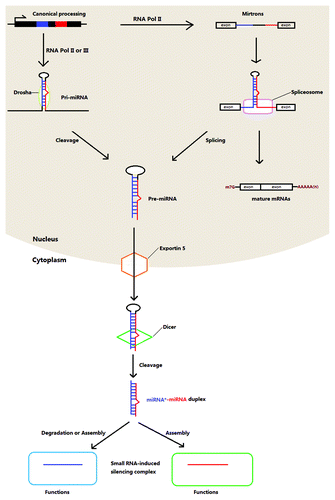
By contrast, the biogenesis of mirtron miRNAs, which are found in C. elegans, Drosophila melanogster,Citation15 chickenCitation18 and several mammals,Citation19 is different. The short intron-derived pri-miRNAs are spliced into intron-removed mRNAs and intron lariats. Without the cleavage by Drosha, the introns are folded directly to form pre-miRNAs with the hairpin structureCitation15 and further processed as described above. Murine herpesvirus miRNAs are produced in yet another way.Citation20 RNA Pol III, instead of Pol II, is functional to produce pri-miRNAs that contain a tRNA-like structure at the 5′ end and two stem loops 1 in the middle and 2 at the 3′ end. The pri-miRNAs are cleaved by tRNase Z but not Drosha to yield the pre-miRNAs that are then further edited in a typical way to liberate mature viral miRNAs.Citation21 In Giardia lamblia, an ancient parasitic protozoan, miRNAs are derived from small nucleolar RNAs (snoRNAs) that participate in modification of other types of RNA.Citation22 Together with other facts that Drosha and Exportin 5 homologs are absent but two vital RISC factors, Argonaute and Dicer, do exist in Giardia raise the possibility of another Drosha-independent pathway is involved in miRNA biogenesis.
The biogenesis of miRNAs from transcription to mature miRNAs is tightly controlled via modifications of miRNA-producing RNA and factors that form functional RISC or other complexes and these modifications can be offered by internal or even external molecules.Citation23 Together with other proteins, such as TRBP and PACT, Dicer constitutes a machinery to process pre-miRNAs and its elevating level can result from the stabilized TRBP through phosphorylation.Citation24 In infected T cells, the abundance of mature miR-27 was alternatively downregulated by binding to one of non-coding RNAs of Herpesvirus saimiri, HSUR1, although the consequent effects of induced decay have not been determined.Citation25
miRNAs in parasite kingdom
At present, miRNAs have been computationally or experimentally investigated in seven protozoans, three trematodes, two nematodes, one cestode and three arthropods (). The lack of coding genes for Dicer and Argonaute proteins, key factors of RISC, in the genome of a number of protozoan parasites, implies the absence of miRNA-induced regulation of gene expression.Citation3 Alignment analysis of Argonaute showed that the expression modulation by small RNA is extensively present across parasitic cestodes and nematodes (). Interestingly, apart from active Argonaute-coding genes, there are Argonaute pseudogenes present in three Leishmania species (), suggesting the shrinking of RNA-inducing silencing functions during evolution. Conversely, the expansion of miRNA pathways has been demonstrated in the plant parasite, Acyrthosiphon pisum, although the biological significance of the miRNA machinery gene duplication events is elusive.Citation26
Table 1. miRNAs and/or RNA-induced silencing machinery in parasites
The loss of this mechanism should be evaluated cautiously; however, in that these coding genes may have undergone great variations in domain(s) during evolution, leading to low similarity and difficulties in identification of putative protein homologs. For instance, Trypanosoma cruzi, the causative pathogen of Chagas disease, was thought to be unable to utilize siRNAs to alter gene expression;Citation3 more recently, however, the presence of a ubiquitously expressed Argonaute/Piwi protein-coding gene has been described.Citation27 Although no potential miRNAs have been found in a small RNA subpopulation,Citation28 it is still hard to rule out the possibility of that this protozoan parasite has the ability to produce miRNAs because of limitations of the methods used.
Characteristics of protozoan miRNA-induced silencing network
Growing studies have demonstrated that miRNA-related pathways in protozoan parasites have features different to other metazoan organisms, possibly shedding light on the evolution of miRNA-induced silencing networks. The RISC components are more heterogeneous in sequence, compared with other animals. Protozoan Argonaute- and Piwi-like genes form an individual lineage, distinct from other Argonaute-like subfamily and Piwi-like subfamily of other multicellular organisms.Citation27 Similarly, phylogenetic analysis of Dicer and RNA-dependent RNA polymerase homologs of Toxoplasma gondii that play a role in magnifying RNA silencing have shown common origins with Chlamydomonas reinhardtii, a unicellular alga that has a capacity of orchestrating gene expression using miRNAs,Citation29 and the fungus Neurospora crassa, respectively.Citation30 It can be partly explained by the concept that the ancestor of apicomplexan animals is proposed to be an endosymbiont of red alga.Citation30 These evidences reinforce the idea that the appearance of the RNA-induced silencing machinery is a primitive event.
Extensive alignment analysis reveals that parasitic protozoans lack Drosha homologs (), indicating that most miRNA biogenesis in these organisms is independent of Drosha. Alternatively, other factor(s) that are significantly divergent in the sequence execute the catalytic activity to liberate pre-miRNAs from primary transcripts. Without the functions of Drosha and Exportin 5, G. lamblia bypasses the canonical Drosha-dependent way to generate miRNAs using snoRNAs as precursors.Citation22
In the target recognition, animal miRNAs are partly complementary to sites of mRNA, majority of which are localized at 3′ UTRs, while plant miRNAs show near-full or full pairing. It has been shown that the extensive base pairing in animal miRNAs induces the turnover of bound small RNA by tailing or trimming.Citation31 In plants, the modification of 2’-O-methyl at the 3′ last nucleotide of miRNAs and siRNAs by Hen1, a methyltransferase, allows these silencing molecules to be immune to uridylation-elicited degradation. The 2’-O-methyl group addition is also required to form the active siRNA-argonaute complex in flies.Citation31 Of particular interest, it was predicted that Toxoplasma miRNAs bind to target regions via nearly perfect or perfect matching.Citation30 In addition to this, more than half of the target sites (20/34) are distributed at 5′ UTRs or in coding regions. Nevertheless, miRNAs are not methylated in T. gondii and the lack of methylation raises the question of how these miRNA are protected. Although the resultant effects are controversial, adenylation at 3′ ends of miRNAs may be a feasible approach adopted to enhance miRNA stabilization.Citation32 Consistent with this, the addition of untemplated adenine found in some Toxoplasma miRNAs render the possibility that miRNAs that are perfectly paired with targets function normally although 3′ adenylation may have an adverse effect on loading miRNA into RISC.Citation33 This may be reflected by the components of the RNA-silencing machinery in T. gondii, of which Dicer does not possess typical DSRM and PAZ domains that associate with RNA binding, are remarkably distinct in structures.
snoRNAs: Original precursors of miRNAs?
snoRNAs are comprised of two types, box C/D and box H/ACA snoRNAs, and combine with other specific factors to be able to modify other types of RNA or function in splicing. In human, both classes of snoRNAs can be a source of small regulatory RNAs (sd-RNAs) that are functionally similar to miRNAs.Citation34,Citation35 Nonetheless, C/D sd-RNAs are distinguished from these out of H/ACA snoRNAs in terms of length and location.Citation36 Conservation of most snoRNAs that can be processed to generate miRNA-like molecules is observed across evolutionarily unrelated species.Citation34,Citation36 sd-RNAs are extensively present in many organisms, including plants, virus and unicellular eukaryotes.Citation36,Citation37 The production of miRNA-like sd-RNAs is Drosha-independent but does require Dicer. Moreover, it has been demonstrated that the capacity of small RNA production is a common characteristic of a portion of snoRNAs.Citation34,Citation38 The extensive discovery that sd-RNAs modulate gene expression in a miRNA-like pattern adds the complexity of RNA-induced silencing mechanism reservoir.
Though the classification of sd-RNAs as miRNAs is unresolved, sd-RNAs and miRNAs are evolutionarily related. snoRNAs and precursors of miRNAs have similar genomic locations. A large number of human snoRNAs reside in introns of genes as well as transposable elements, from which many miRNAs are derived.Citation39,Citation40 Moreover, both types of small silencing molecules exhibit conservation throughout evolution.Citation34,Citation36,Citation37 Furthermore, like miRNAs, upon loading into Argonaute-containing RISC, sd-RNAs induce gene silencing via Watson-Crick base pairing to target sites of mRNA they regulate. Not surprisingly, some registered human miRNAs (miRBase) are really derived from snoRNAs.Citation38,Citation41 Of note, some sd-RNAs are expressed in a specific cell-type manner, likely due to the post-transcriptional process of snoRNAs as do some miRNA precursors.Citation38,Citation42 However, the second structures of snoRNAs contain two hairpins, obviously different from classical miRNA precursors. Taken together suggests the evolutionary relationship between snoRNAs and miRNA pathways.
Functional similarity of these silencing RNAs is supported by the interactions between Argonaute proteins and some core components of snoRNPs, such as NOP56.Citation30,Citation43 Although the real functions are unclear, the presence of NOP56 in RISC may reveal that sd-RNAs are expressed in T. gondii. The fact that miRNAs are exclusively derived from snoRNAs in unicellular animal, G. lamblia, results in the hypothesis that snoRNAs are the original source of miRNAs.Citation22 In agreement with this, a portion of miRNA precursors did have box H/ACA snoRNA properties and all five snoRNA-like miRNA precursors investigated showed capacity of binding with dyskerin,Citation41 a pseudouridine synthase in snoRNA-containing nucleolar ribonucleoparticles (snoRNPs),Citation44 suggesting the preservation of snoRNA functions in these miRNA precursors. snoRNAs being derived from transposable elements as miRNA precursors may be likely during evolution because the rapid generation of miRNAs through transposable elements could be a driving force.Citation39 It is no doubt that the clarification of how snoRNAs are processed into sd-RNAs allows us to profoundly understand the relationship between sd-RNAs and miRNAs.
snoRNAs are also reported in Plasmodium falciparum,Citation45,Citation46 Trypanosoma spp,Citation47,Citation48 Trichomonas vaginalisCitation49 and S. mansoni and S. japonicumCitation50 but their ability to encode miRNA-like sd-RNAs remains unclear. It is noticed that P. falciparum and Saccharomyces cervisiae express snoRNAs but no miRNAs, in agreement with the idea that primitive snoRNAs may have produced certain types of miRNA-like RNA.Citation41
Potential miRNA biomarkers for flatworm neoblasts
In the phylum Platyhelminthes, planarians are characterized by the ability to regenerateCitation51 and neoblasts are thought to contribute to this regeneration. The neoblasts of E. multilocularis are able to generate mature metacestodes in vitro under specific conditions,Citation52 suggesting the preservation of regenerative capacity in the neoblasts.
Fifteen miRNAs, including let-7a and 7b, have been implicated in the functions of the planaria neoblasts,Citation51,Citation53-Citation55 although there is some disparity probably due to methodological differences. Aligned with planarian miRNAs, there are six potential miRNA neoblast markers commonly found in S. japonicum and E. multilocularis, in all of which the seed regions are intact (). Interestingly, miR-71 with other three miRNAs miR-2d, miR-752 and miR-13 are clustered in planarians but miR-752 is completely lost in S. japonicum and E. multilocularis. Moreover, two copies of the miR71 clusters are observed in S. japonicumCitation56 but not in E. multilocularis (our unpublished data), suggesting duplication of miR71 cluster after speciation. What the loss of miR-752 influences the regenerative ability of neoblasts is unknown and remains to be identified experimentally.
Table 2. Potential neoblast-specific miRNAs
Possible roles of parasite miRNAs
An exactly spatial and temporal control of gene expression is crucial for animals whose life cycles are fulfilled through several different developmental stages. microRNAs, such as the let-7 family and lin-4, were first revealed to be a master switch of development in C. elegans,Citation11 targeting hb1-7 and lin-14 mRNAs, respectively, to ensure proper larval developmental transition. The transcription of let-7 family is controlled by the interaction between the nuclear hormone receptor DAF-12 and its ligands, dafachronic acids that are naturally produced in favorable environments.Citation57 The striking discrepancy of expression of Dicer and Argonaute at different stages in S. mansoni is an indication that small RNA-induced silencing represents a mechanism of modulation of developmental transitions.Citation58 The idea that S. japonicum let-7 may associate with transition from miracidium to sporocyst is indirectly supported by higher expression of let-7 in miracidium. Along with let-7, the miRNA bantam that is extraordinarily highly expressed in cercaria is supposed to be active in developmental processes through regulation of a cellular population size.Citation59 However, a recent study preferably supports the notion that let-7 has other functions rather than controlling developmental timing.Citation60 It is clear that these presumptions need to be further experimentally tested.
Some miRNAs are anticipated to take part in other processes of parasites, such as reproduction and tissue development (). Of interest is the very high expression of miR-71(a/b) in stages in trematodesCitation56,Citation59,Citation60 and a cestode (our unpublished data) but not in a nematode.Citation61 Apart from functional connections with neoblast biology,Citation53-Citation55 miR-71 is expected to have an additional role probably related to development. Further experiments will be worthwhile specifying the connections between its expression and functions.
Table 3. Potential functions of parasite miRNAs
miRNAs are modulators in immune systems, affecting differentiation, development, homeostasis and functions of immune cells.Citation62,Citation63 It is well-studied that miRNAs play important roles in viral infection and immunology.Citation25,Citation64-Citation67 The temporal regulation of variant surface proteins (VSGs), one of which covers the entire surface of a parasite at any time, is essential to survive host immune attacks in free living protozoans such as Trypanosomes and G. lamblia.Citation68,Citation69 Various VSGs expressed on the surface were observed in individual G. lamblia with silenced Dicer and RNA-dependent RNA polymerase, indicating the key role of silencing machinery in VSG expression regulation.Citation14 miRNAs may be partly responsible for this antigenic variation by targeting 3′ VSG-coding gene UTRs that contain highly conserved fragments.Citation22,Citation68 The mechanism of how miRNA(s) modulate several hundreds of VSG genes to ensure that only one is retained is unknown.
Unfortunately, little has been done to experimentally investigate the functions of parasite miRNAs but a wealth of the genome, transcriptome and proteome data will accelerate to characterize their roles in parasites and parasitic infection.
Host miRNAs in response to parasite infection
The alterations of host miRNA expression reflect the roles of host-derived miRNAs in parasite infection. Cryptosporidium parvum, a protozoan parasite which lacks the RNA-induced silencing mechanism (), can induce a decrease of let-7 expression in infected cells, leading to upregulation of Toll-like receptor 4 (TLR4) expression, which contains a let-7 binding site in the 3′ UTR.Citation70 In vitro suppression of let-7 gives rise to a significantly lower parasite burden, suggesting that it contributes to immune responses against the infection. The parasite-induced downregulation of let-7 expression is executed by a repressor binding to the Let-7 promoter, which is comprised of transcription factors NFκB p50 and CCAAT/enhancer-binding protein β.Citation71 By contrast, a number of miRNAs was upregulated in a NFκB p65-dependent or -independent manner in response to C. parvum invasion ().Citation72 Functional silencing of selected miRNAs resulted in an increase of C. parvum burden in vitro without effects on cell attachment and invasion, suggesting a role of host miRNAs in epithelial defense against parasite infection.
Figure 2. Host miRNAs response to Cryptosporidium infection. During the period of infection, Cryptosporidum can elicit alterations of host-origin miRNA expression. Some miRNAs are repressed (in blue), whereas others upregulated (in red) in NFκB p65-dependent or -independent (?) manner. Protein levels of some key players involved in infection and immunology will be subsequently increased (in red), facilitating parasite clearance or infection.
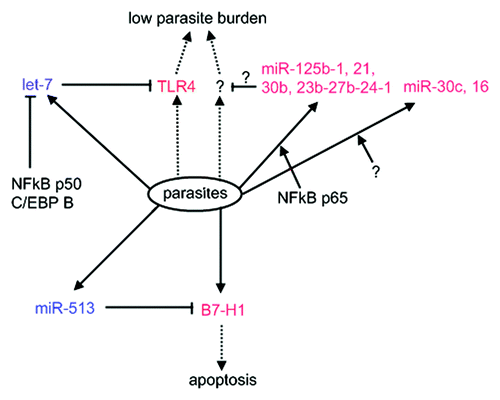
C. parvum can repress miR-513 transcription and simultaneously activate the expression of B7-H1, residing a binding site for miR-513. This leads to an increase of B7-H1expressed on the cell surface that in vitro induces the apoptosis of activated T cells.Citation73 This is of particular interest because it illustrates the feasibility that parasites take advantage of host miRNA pathways as a method of defense against the host. The in vivo events following increased B7-H1 will be of interest to be further investigated. Parasite-induced host miRNA expression alterations were also observed in T. gondii, though the relationship between elevating miRNA level and the infection is not clear.Citation74 In contrast to the unaltered level in response to Neospora caninum infection, the expression of mature miR-17 family was upregulated in the cells infected by T. gondii, illustrating the elevated abundance specific to T. gondii infection. Collectively, host miRNA-induced silencing networks take part in interplay between host and pathogen via modulation of key players that are active in a course of infection and immunology.
Concluding Remarks
Although the vast majority of miRNAs are enigmatic in function, it is clear that the miRNA-induced silencing machinery directly or indirectly affects many processes of organisms and their responses to environments. Parasite and host miRNA profiles can be served as a probe to investigate underlying mechanisms and, thus, deeply understand pathogen-host interplay. Likewise, miRNA silencing network will be an alternative to help us understand global deleterious drug resistance in parasitic nematodes.Citation75 The intervention of miRNA pathways to control diseases is in infancy, but successful attempts to control viral infection in chimpanzees or tumorigenesis in animal modelsCitation76,Citation77 have shed light on the potential of miRNA pathways as the therapeutical targets.
Acknowledgments
This study was financially supported by National Natural Science Foundation of China (31201900), State Public-interest Institution Basal Research Fund, Chinese Academy of Agricultural Sciences (0032012037), the Open Fund of the Key Laboratory of the New Animal Drug Project of Gansu Province and the Key Laboratory of Veterinary Pharmaceutical Development of the Ministry of Agriculture (1610322011011) and by the Science Fund for Creative Research Groups of Gansu Province (Grant No.1210RJIA006). The part of analyses in this study was conducted in the University of Nottingham under the financial support of Overseas Research Students Awards (Y.Z.), UK and the University of Nottingham scholarship (Y.Z.). The authors would like to give thanks to reviewers for their constructive suggestions. We also thank institutes or sponsors for free use of unpublished genomic data.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Related Research Data
References
- Grimson A, Srivastava M, Fahey B, Woodcroft BJ, Chiang HR, King N, et al. Early origins and evolution of microRNAs and Piwi-interacting RNAs in animals. Nature 2008; 455:1193 - 7; http://dx.doi.org/10.1038/nature07415; PMID: 18830242
- Rathjen T, Nicol C, McConkey G, Dalmay T. Analysis of short RNAs in the malaria parasite and its red blood cell host. FEBS Lett 2006; 580:5185 - 8; http://dx.doi.org/10.1016/j.febslet.2006.08.063; PMID: 16963026
- Militello KT, Refour P, Comeaux CA, Duraisingh MT. Antisense RNA and RNAi in protozoan parasites: working hard or hardly working?. Mol Biochem Parasitol 2008; 157:117 - 26; http://dx.doi.org/10.1016/j.molbiopara.2007.10.004; PMID: 18053590
- Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008; 455:58 - 63; http://dx.doi.org/10.1038/nature07228; PMID: 18668040
- Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP. The impact of microRNAs on protein output. Nature 2008; 455:64 - 71; http://dx.doi.org/10.1038/nature07242; PMID: 18668037
- Xie X, Lu J, Kulbokas EJ, Golub TR, Mootha V, Lindblad-Toh K, et al. Systematic discovery of regulatory motifs in human promoters and 3′ UTRs by comparison of several mammals. Nature 2005; 434:338 - 45; http://dx.doi.org/10.1038/nature03441; PMID: 15735639
- Miska EA, Alvarez-Saavedra E, Abbott AL, Lau NC, Hellman AB, McGonagle SM, et al. Most Caenorhabditis elegans microRNAs are individually not essential for development or viability. PLoS Genet 2007; 3:e215; http://dx.doi.org/10.1371/journal.pgen.0030215; PMID: 18085825
- Li X, Cassidy JJ, Reinke CA, Fischboeck S, Carthew RW. A microRNA imparts robustness against environmental fluctuation during development. Cell 2009; 137:273 - 82; http://dx.doi.org/10.1016/j.cell.2009.01.058; PMID: 19379693
- Lewis MA, Quint E, Glazier AM, Fuchs H, De Angelis MH, Langford C, et al. An ENU-induced mutation of miR-96 associated with progressive hearing loss in mice. Nat Genet 2009; 41:614 - 8; http://dx.doi.org/10.1038/ng.369; PMID: 19363478
- He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, Goodson S, et al. A microRNA polycistron as a potential human oncogene. Nature 2005; 435:828 - 33; http://dx.doi.org/10.1038/nature03552; PMID: 15944707
- Kloosterman WP, Plasterk RH. The diverse functions of microRNAs in animal development and disease. Dev Cell 2006; 11:441 - 50; http://dx.doi.org/10.1016/j.devcel.2006.09.009; PMID: 17011485
- Ma L, Weinberg RA. Micromanagers of malignancy: role of microRNAs in regulating metastasis. Trends Genet 2008; 24:448 - 56; http://dx.doi.org/10.1016/j.tig.2008.06.004; PMID: 18674843
- Winter F, Edaye S, Hüttenhofer A, Brunel C. Anopheles gambiae miRNAs as actors of defence reaction against Plasmodium invasion.. Nucleic Acids Res 2007; 35:6953 - 62; http://dx.doi.org/10.1093/nar/gkm686; PMID: 17933784
- Prucca CG, Slavin I, Quiroga R, Elías EV, Rivero FD, Saura A, et al. Antigenic variation in Giardia lamblia is regulated by RNA interference. Nature 2008; 456:750 - 4; http://dx.doi.org/10.1038/nature07585; PMID: 19079052
- Ruby JG, Jan CH, Bartel DP. Intronic microRNA precursors that bypass Drosha processing. Nature 2007; 448:83 - 6; http://dx.doi.org/10.1038/nature05983; PMID: 17589500
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004; 116:281 - 97; http://dx.doi.org/10.1016/S0092-8674(04)00045-5; PMID: 14744438
- Okamura K, Phillips MD, Tyler DM, Duan H, Chou YT, Lai EC. The regulatory activity of microRNA* species has substantial influence on microRNA and 3′ UTR evolution. Nat Struct Mol Biol 2008; 15:354 - 63; http://dx.doi.org/10.1038/nsmb.1409; PMID: 18376413
- Glazov EA, Cottee PA, Barris WC, Moore RJ, Dalrymple BP, Tizard ML. A microRNA catalog of the developing chicken embryo identified by a deep sequencing approach. Genome Res 2008; 18:957 - 64; http://dx.doi.org/10.1101/gr.074740.107; PMID: 18469162
- Berezikov E, Chung WJ, Willis J, Cuppen E, Lai EC. Mammalian mirtron genes. Mol Cell 2007; 28:328 - 36; http://dx.doi.org/10.1016/j.molcel.2007.09.028; PMID: 17964270
- Bogerd HP, Karnowski HW, Cai X, Shin J, Pohlers M, Cullen BR. A mammalian herpesvirus uses noncanonical expression and processing mechanisms to generate viral MicroRNAs. Mol Cell 2010; 37:135 - 42; http://dx.doi.org/10.1016/j.molcel.2009.12.016; PMID: 20129062
- Scherer LJ, Frank R, Rossi JJ. Optimization and characterization of tRNA-shRNA expression constructs. Nucleic Acids Res 2007; 35:2620 - 8; http://dx.doi.org/10.1093/nar/gkm103; PMID: 17426139
- Saraiya AA, Wang CC. snoRNA, a novel precursor of microRNA in Giardia lamblia.. PLoS Pathog 2008; 4:e1000224; http://dx.doi.org/10.1371/journal.ppat.1000224; PMID: 19043559
- Kim YK, Heo I, Kim VN. Modifications of small RNAs and their associated proteins. Cell 2010; 143:703 - 9; http://dx.doi.org/10.1016/j.cell.2010.11.018; PMID: 21111232
- Paroo Z, Ye X, Chen S, Liu Q. Phosphorylation of the human microRNA-generating complex mediates MAPK/Erk signaling. Cell 2009; 139:112 - 22; http://dx.doi.org/10.1016/j.cell.2009.06.044; PMID: 19804757
- Cazalla D, Yario T, Steitz JA. Down-regulation of a host microRNA by a Herpesvirus saimiri noncoding RNA. Science 2010; 328:1563 - 6; http://dx.doi.org/10.1126/science.1187197; PMID: 20558719
- Jaubert-Possamai S, Rispe C, Tanguy S, Gordon K, Walsh T, Edwards O, et al. Expansion of the miRNA pathway in the hemipteran insect Acyrthosiphon pisum.. Mol Biol Evol 2010; 27:979 - 87; http://dx.doi.org/10.1093/molbev/msp256; PMID: 20179251
- Garcia Silva MR, Tosar JP, Frugier M, Pantano S, Bonilla B, Esteban L, et al. Cloning, characterization and subcellular localization of a Trypanosoma cruzi argonaute protein defining a new subfamily distinctive of trypanosomatids. Gene 2010; 466:26 - 35; http://dx.doi.org/10.1016/j.gene.2010.06.012; PMID: 20621168
- Garcia-Silva MR, Frugier M, Tosar JP, Correa-Dominguez A, Ronalte-Alves L, Parodi-Talice A, et al. A population of tRNA-derived small RNAs is actively produced in Trypanosoma cruzi and recruited to specific cytoplasmic granules. Mol Biochem Parasitol 2010; 171:64 - 73; http://dx.doi.org/10.1016/j.molbiopara.2010.02.003; PMID: 20156490
- Molnár A, Schwach F, Studholme DJ, Thuenemann EC, Baulcombe DC. miRNAs control gene expression in the single-cell alga Chlamydomonas reinhardtii.. Nature 2007; 447:1126 - 9; http://dx.doi.org/10.1038/nature05903; PMID: 17538623
- Braun L, Cannella D, Ortet P, Barakat M, Sautel CF, Kieffer S, et al. A complex small RNA repertoire is generated by a plant/fungal-like machinery and effected by a metazoan-like Argonaute in the single-cell human parasite Toxoplasma gondii.. PLoS Pathog 2010; 6:e1000920; http://dx.doi.org/10.1371/journal.ppat.1000920; PMID: 20523899
- Ameres SL, Horwich MD, Hung JH, Xu J, Ghildiyal M, Weng Z, et al. Target RNA-directed trimming and tailing of small silencing RNAs. Science 2010; 328:1534 - 9; http://dx.doi.org/10.1126/science.1187058; PMID: 20558712
- Lu S, Sun YH, Chiang VL. Adenylation of plant miRNAs. Nucleic Acids Res 2009; 37:1878 - 85; http://dx.doi.org/10.1093/nar/gkp031; PMID: 19188256
- Burroughs AM, Ando Y, de Hoon MJ, Tomaru Y, Nishibu T, Ukekawa R, et al. A comprehensive survey of 3′ animal miRNA modification events and a possible role for 3′ adenylation in modulating miRNA targeting effectiveness. Genome Res 2010; 20:1398 - 410; http://dx.doi.org/10.1101/gr.106054.110; PMID: 20719920
- Ender C, Krek A, Friedländer MR, Beitzinger M, Weinmann L, Chen W, et al. A human snoRNA with microRNA-like functions. Mol Cell 2008; 32:519 - 28; http://dx.doi.org/10.1016/j.molcel.2008.10.017; PMID: 19026782
- Brameier M, Herwig A, Reinhardt R, Walter L, Gruber J. Human box C/D snoRNAs with miRNA like functions: expanding the range of regulatory RNAs. Nucleic Acids Res 2011; 39:675 - 86; http://dx.doi.org/10.1093/nar/gkq776; PMID: 20846955
- Taft RJ, Glazov EA, Lassmann T, Hayashizaki Y, Carninci P, Mattick JS. Small RNAs derived from snoRNAs. RNA 2009; 15:1233 - 40; http://dx.doi.org/10.1261/rna.1528909; PMID: 19474147
- Hutzinger R, Feederle R, Mrazek J, Schiefermeier N, Balwierz PJ, Zavolan M, et al. Expression and processing of a small nucleolar RNA from the Epstein-Barr virus genome. PLoS Pathog 2009; 5:e1000547; http://dx.doi.org/10.1371/journal.ppat.1000547; PMID: 19680535
- Brameier M, Herwig A, Reinhardt R, Walter L, Gruber J. Human box C/D snoRNAs with miRNA like functions: expanding the range of regulatory RNAs. Nucleic Acids Res 2011; 39:675 - 86; http://dx.doi.org/10.1093/nar/gkq776; PMID: 20846955
- Smalheiser NR, Torvik VI. Mammalian microRNAs derived from genomic repeats. Trends Genet 2005; 21:322 - 6; http://dx.doi.org/10.1016/j.tig.2005.04.008; PMID: 15922829
- Piriyapongsa J, Mariño-Ramírez L, Jordan IK. Origin and evolution of human microRNAs from transposable elements. Genetics 2007; 176:1323 - 37; http://dx.doi.org/10.1534/genetics.107.072553; PMID: 17435244
- Scott MS, Avolio F, Ono M, Lamond AI, Barton GJ. Human miRNA precursors with box H/ACA snoRNA features. PLoS Comput Biol 2009; 5:e1000507; http://dx.doi.org/10.1371/journal.pcbi.1000507; PMID: 19763159
- Obernosterer G, Leuschner PJ, Alenius M, Martinez J. Post-transcriptional regulation of microRNA expression. RNA 2006; 12:1161 - 7; http://dx.doi.org/10.1261/rna.2322506; PMID: 16738409
- Höck J, Weinmann L, Ender C, Rüdel S, Kremmer E, Raabe M, et al. Proteomic and functional analysis of Argonaute-containing mRNA-protein complexes in human cells. EMBO Rep 2007; 8:1052 - 60; http://dx.doi.org/10.1038/sj.embor.7401088; PMID: 17932509
- Hoang C, Ferré-D’Amaré AR. Cocrystal structure of a tRNA Psi55 pseudouridine synthase: nucleotide flipping by an RNA-modifying enzyme. Cell 2001; 107:929 - 39; http://dx.doi.org/10.1016/S0092-8674(01)00618-3; PMID: 11779468
- Raabe CA, Sanchez CP, Randau G, Robeck T, Skryabin BV, Chinni SV, et al. A global view of the nonprotein-coding transcriptome in Plasmodium falciparum.. Nucleic Acids Res 2010; 38:608 - 17; http://dx.doi.org/10.1093/nar/gkp895; PMID: 19864253
- Chakrabarti K, Pearson M, Grate L, Sterne-Weiler T, Deans J, Donohue JP, et al. Structural RNAs of known and unknown function identified in malaria parasites by comparative genomics and RNA analysis. RNA 2007; 13:1923 - 39; http://dx.doi.org/10.1261/rna.751807; PMID: 17901154
- Uliel S, Liang XH, Unger R, Michaeli S. Small nucleolar RNAs that guide modification in trypanosomatids: repertoire, targets, genome organisation, and unique functions. Int J Parasitol 2004; 34:445 - 54; http://dx.doi.org/10.1016/j.ijpara.2003.10.014; PMID: 15013734
- Morales L, Romero I, Diez H, Del Portillo P, Montilla M, Nicholls S, et al. Characterization of a candidate Trypanosoma rangeli small nucleolar RNA gene and its application in a PCR-based parasite detection. Exp Parasitol 2002; 102:72 - 80; http://dx.doi.org/10.1016/S0014-4894(03)00027-4; PMID: 12706742
- Chen XS, Collins LJ, Biggs PJ, Penny D. High throughput genome-wide survey of small RNAs from the parasitic protists Giardia intestinalis and Trichomonas vaginalis.. Genome Biol Evol 2009; 1:165 - 75; http://dx.doi.org/10.1093/gbe/evp017; PMID: 20333187
- Copeland CS, Marz M, Rose D, Hertel J, Brindley PJ, Santana CB, et al. Homology-based annotation of non-coding RNAs in the genomes of Schistosoma mansoni and Schistosoma japonicum.. BMC Genomics 2009; 10:464; http://dx.doi.org/10.1186/1471-2164-10-464; PMID: 19814823
- Palakodeti D, Smielewska M, Graveley BR. MicroRNAs from the Planarian Schmidtea mediterranea: a model system for stem cell biology. RNA 2006; 12:1640 - 9; http://dx.doi.org/10.1261/rna.117206; PMID: 16849698
- Spiliotis M, Lechner S, Tappe D, Scheller C, Krohne G, Brehm K. Transient transfection of Echinococcus multilocularis primary cells and complete in vitro regeneration of metacestode vesicles. Int J Parasitol 2008; 38:1025 - 39; http://dx.doi.org/10.1016/j.ijpara.2007.11.002; PMID: 18086473
- Lu YC, Smielewska M, Palakodeti D, Lovci MT, Aigner S, Yeo GW, et al. Deep sequencing identifies new and regulated microRNAs in Schmidtea mediterranea.. RNA 2009; 15:1483 - 91; http://dx.doi.org/10.1261/rna.1702009; PMID: 19553344
- Friedländer MR, Adamidi C, Han T, Lebedeva S, Isenbarger TA, Hirst M, et al. High-resolution profiling and discovery of planarian small RNAs. Proc Natl Acad Sci USA 2009; 106:11546 - 51; http://dx.doi.org/10.1073/pnas.0905222106; PMID: 19564616
- González-Estévez C, Arseni V, Thambyrajah RS, Felix DA, Aboobaker AA. Diverse miRNA spatial expression patterns suggest important roles in homeostasis and regeneration in planarians. Int J Dev Biol 2009; 53:493 - 505; http://dx.doi.org/10.1387/ijdb.082825cg; PMID: 19247960
- Huang J, Hao P, Chen H, Hu W, Yan Q, Liu F, et al. Genome-wide identification of Schistosoma japonicum microRNAs using a deep-sequencing approach. PLoS One 2009; 4:e8206; http://dx.doi.org/10.1371/journal.pone.0008206; PMID: 19997615
- Bethke A, Fielenbach N, Wang Z, Mangelsdorf DJ, Antebi A. Nuclear hormone receptor regulation of microRNAs controls developmental progression. Science 2009; 324:95 - 8; http://dx.doi.org/10.1126/science.1164899; PMID: 19342589
- Gomes MS, Cabral FJ, Jannotti-Passos LK, Carvalho O, Rodrigues V, Baba EH, et al. Preliminary analysis of miRNA pathway in Schistosoma mansoni.. Parasitol Int 2009; 58:61 - 8; http://dx.doi.org/10.1016/j.parint.2008.10.002; PMID: 19007911
- Xue X, Sun J, Zhang Q, Wang Z, Huang Y, Pan W. Identification and characterization of novel microRNAs from Schistosoma japonicum.. PLoS One 2008; 3:e4034; http://dx.doi.org/10.1371/journal.pone.0004034; PMID: 19107204
- Hao L, Cai P, Jiang N, Wang H, Chen Q. Identification and characterization of microRNAs and endogenous siRNAs in Schistosoma japonicum.. BMC Genomics 2010; 11:55; http://dx.doi.org/10.1186/1471-2164-11-55; PMID: 20092619
- Huang QX, Cheng XY, Mao ZC, Wang YS, Zhao LL, Yan X, et al. MicroRNA discovery and analysis of pinewood nematode Bursaphelenchus xylophilus by deep sequencing. PLoS One 2010; 5:e13271; http://dx.doi.org/10.1371/journal.pone.0013271; PMID: 20967259
- Xiao C, Rajewsky K. MicroRNA control in the immune system: basic principles. Cell 2009; 136:26 - 36; http://dx.doi.org/10.1016/j.cell.2008.12.027; PMID: 19135886
- Baltimore D, Boldin MP, O’Connell RM, Rao DS, Taganov KD. MicroRNAs: new regulators of immune cell development and function. Nat Immunol 2008; 9:839 - 45; http://dx.doi.org/10.1038/ni.f.209; PMID: 18645592
- Triboulet R, Mari B, Lin YL, Chable-Bessia C, Bennasser Y, Lebrigand K, et al. Suppression of microRNA-silencing pathway by HIV-1 during virus replication. Science 2007; 315:1579 - 82; http://dx.doi.org/10.1126/science.1136319; PMID: 17322031
- Jopling CL, Yi M, Lancaster AM, Lemon SM, Sarnow P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science 2005; 309:1577 - 81; http://dx.doi.org/10.1126/science.1113329; PMID: 16141076
- Nachmani D, Stern-Ginossar N, Sarid R, Mandelboim O. Diverse herpesvirus microRNAs target the stress-induced immune ligand MICB to escape recognition by natural killer cells. Cell Host Microbe 2009; 5:376 - 85; http://dx.doi.org/10.1016/j.chom.2009.03.003; PMID: 19380116
- Ghosh Z, Mallick B, Chakrabarti J. Cellular versus viral microRNAs in host-virus interaction. Nucleic Acids Res 2009; 37:1035 - 48; http://dx.doi.org/10.1093/nar/gkn1004; PMID: 19095692
- Nash TE. Surface antigenic variation in Giardia lamblia.. Mol Microbiol 2002; 45:585 - 90; http://dx.doi.org/10.1046/j.1365-2958.2002.03029.x; PMID: 12139606
- Mallick B, Ghosh Z, Chakrabarti J. MicroRNA switches in Trypanosoma brucei.. Biochem Biophys Res Commun 2008; 372:459 - 63; http://dx.doi.org/10.1016/j.bbrc.2008.05.084; PMID: 18510949
- Chen XM, Splinter PL, O’Hara SP, LaRusso NF. A cellular micro-RNA, let-7i, regulates Toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. J Biol Chem 2007; 282:28929 - 38; http://dx.doi.org/10.1074/jbc.M702633200; PMID: 17660297
- O’Hara SP, Splinter PL, Gajdos GB, Trussoni CE, Fernandez-Zapico ME, Chen XM, et al. NFkappaB p50-CCAAT/enhancer-binding protein beta (C/EBPbeta)-mediated transcriptional repression of microRNA let-7i following microbial infection. J Biol Chem 2010; 285:216 - 25; http://dx.doi.org/10.1074/jbc.M109.041640; PMID: 19903813
- Zhou R, Hu G, Liu J, Gong AY, Drescher KM, Chen XM. NF-kappaB p65-dependent transactivation of miRNA genes following Cryptosporidium parvum infection stimulates epithelial cell immune responses. PLoS Pathog 2009; 5:e1000681; http://dx.doi.org/10.1371/journal.ppat.1000681; PMID: 19997496
- Gong AY, Zhou R, Hu G, Liu J, Sosnowska D, Drescher KM, et al. Cryptosporidium parvum induces B7-H1 expression in cholangiocytes by down-regulating microRNA-513. J Infect Dis 2010; 201:160 - 9; http://dx.doi.org/10.1086/648589; PMID: 19916867
- Zeiner GM, Norman KL, Thomson JM, Hammond SM, Boothroyd JC. Toxoplasma gondii infection specifically increases the levels of key host microRNAs. PLoS One 2010; 5:e8742; http://dx.doi.org/10.1371/journal.pone.0008742; PMID: 20090903
- Devaney E, Winter AD, Britton C. microRNAs: a role in drug resistance in parasitic nematodes?. Trends Parasitol 2010; 26:428 - 33; http://dx.doi.org/10.1016/j.pt.2010.05.003; PMID: 20541972
- Lanford RE, Hildebrandt-Eriksen ES, Petri A, Persson R, Lindow M, Munk ME, et al. Therapeutic silencing of microRNA-122 in primates with chronic hepatitis C virus infection. Science 2010; 327:198 - 201; http://dx.doi.org/10.1126/science.1178178; PMID: 19965718
- Kota J, Chivukula RR, O’Donnell KA, Wentzel EA, Montgomery CL, Hwang HW, et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009; 137:1005 - 17; http://dx.doi.org/10.1016/j.cell.2009.04.021; PMID: 19524505
- De S, Pal D, Ghosh SK. Entamoeba histolytica: computational identification of putative microRNA candidates. Exp Parasitol 2006; 113:239 - 43; http://dx.doi.org/10.1016/j.exppara.2006.01.009; PMID: 16515787
- Xu MJ, Liu Q, Nisbet AJ, Cai XQ, Yan C, Lin RQ, et al. Identification and characterization of microRNAs in Clonorchis sinensis of human health significance. BMC Genomics 2010; 11:521; http://dx.doi.org/10.1186/1471-2164-11-521; PMID: 20920166
- Wang Z, Xue X, Sun J, Luo R, Xu X, Jiang Y, et al. An “in-depth” description of the small non-coding RNA population of Schistosoma japonicum schistosomulum. PLoS Negl Trop Dis 2010; 4:e596; http://dx.doi.org/10.1371/journal.pntd.0000596; PMID: 20161724
- Poole CB, Davis PJ, Jin J, McReynolds LA. Cloning and bioinformatic identification of small RNAs in the filarial nematode, Brugia malayi.. Mol Biochem Parasitol 2010; 169:87 - 94; http://dx.doi.org/10.1016/j.molbiopara.2009.10.004; PMID: 19874857
- Cucher M, Prada L, Mourglia-Ettlin G, Dematteis S, Camicia F, Asurmendi S, et al. Identification of Echinococcus granulosus microRNAs and their expression in different life cycle stages and parasite genotypes. Int J Parasitol 2011; 41:439 - 48; http://dx.doi.org/10.1016/j.ijpara.2010.11.010; PMID: 21219906
- Kirkness EF, Haas BJ, Sun W, Braig HR, Perotti MA, Clark JM, et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc Natl Acad Sci USA 2010; 107:12168 - 73; http://dx.doi.org/10.1073/pnas.1003379107; PMID: 20566863
- International Aphid Genomics Consortium. Genome sequence of the pea aphid Acyrthosiphon pisum.. PLoS Biol 2010; 8:e1000313; http://dx.doi.org/10.1371/journal.pbio.1000313; PMID: 20186266
- Wheeler BM, Heimberg AM, Moy VN, Sperling EA, Holstein TW, Heber S, et al. The deep evolution of metazoan microRNAs. Evol Dev 2009; 11:50 - 68; http://dx.doi.org/10.1111/j.1525-142X.2008.00302.x; PMID: 19196333