
Abstract
During recent years, riboswitches have emerged as potential targets for novel antibacterial substances. In this study, we investigated how one flavin analog, roseoflavin, affected the gene-expression, growth and infectivity of the human bacterial pathogen Listeria monocytogenes to determine the potential of this analog to function as an antibacterial substance. The results indicate that roseoflavin has a profound inhibiting effect on the growth of L. monocytogenes at very low concentrations. Also, expression of the gene located downstream of the FMN riboswitch, a riboflavin transporter, was blocked by the addition of roseoflavin. Base-substitution mutations in the FMN riboswitch allowed the bacteria to grow in the presence of roseoflavin, showing that roseoflavin targeted the FMN riboswitch directly. Surprisingly, we found that roseoflavin stimulated L. monocytogenes virulence gene expression and infection abilities in a mechanism independent of the FMN riboswitch. Our results suggest that roseoflavin can block growth but also enhance Listeria virulence.
Acknowledgements
J.J. is supported by Umeå University, the Swedish Research Council grants (K2011-56X-15144-08-6 and 621-2009-5677) and an ERC starting grant (no 260764-RNAntibiotics).
Figures and Tables
Figure 1 Chemical structures of natural metabolites and analogs. (A) Roseoflavin (B) Riboflavin.
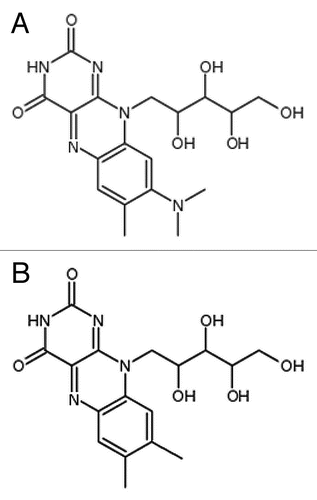
Figure 2 Roseoflavin inhibits the growth of L. monocytogenes and expression of Imo1945 in minimal medium. (A) Bacteria were grown for 10 hours in minimal medium in the absence or presence of roseoflavin (Ro) or riboflavin (Ri) at a concentration of 100 µM. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 4) and statistical analysis [Student t-test (two-tailed) (***p < 0.001)]. (B) Genetic context of the FMN riboswitch Rli96 and the downstream gene Imo1945. Transcription initiates at a promoter generating Rli96. In presence of a functional metabolite binding Rli96, a structural re-arrangement causes transcription termination at the hatched lollipop, generating Rli96 (hatched short line). In absence of such metabolite, transcription proceeds and an Rli96 + Imo1945 transcript is formed (hatched longer line). (C) L. monocytogenes was grown in minimal medium to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to Rli96 and tmRNA (control), respectively.
![Figure 2 Roseoflavin inhibits the growth of L. monocytogenes and expression of Imo1945 in minimal medium. (A) Bacteria were grown for 10 hours in minimal medium in the absence or presence of roseoflavin (Ro) or riboflavin (Ri) at a concentration of 100 µM. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 4) and statistical analysis [Student t-test (two-tailed) (***p < 0.001)]. (B) Genetic context of the FMN riboswitch Rli96 and the downstream gene Imo1945. Transcription initiates at a promoter generating Rli96. In presence of a functional metabolite binding Rli96, a structural re-arrangement causes transcription termination at the hatched lollipop, generating Rli96 (hatched short line). In absence of such metabolite, transcription proceeds and an Rli96 + Imo1945 transcript is formed (hatched longer line). (C) L. monocytogenes was grown in minimal medium to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to Rli96 and tmRNA (control), respectively.](/cms/asset/0c358430-781c-4352-9511-25837c750d02/krnb_a_10915586_f0002.gif)
Figure 3 Roseoflavin inhibits the growth of L. monocytogenes and the expression of Imo1945 in BHI. Bacteria were grown for 10 hours in BHI in the absence or presence of roseoflavin (Ro) or riboflavin (Ri) at a concentration of 100 µM. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 3) and statistical analysis [Student t-test (two-tailed) (***p < 0.001) or (*p < 0.05)]. (B) Bacteria were grown for 10 hours in BHI in the absence or presence of roseoflavin at a concentration of 1, 10 or 100 µM, respectively. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 3) and statistical analysis [Student t-test (two-tailed) (***p < 0.001)]. (C) L. monocytogenes was grown in BHI to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to Rli96 and tmRNA (control), respectively.
![Figure 3 Roseoflavin inhibits the growth of L. monocytogenes and the expression of Imo1945 in BHI. Bacteria were grown for 10 hours in BHI in the absence or presence of roseoflavin (Ro) or riboflavin (Ri) at a concentration of 100 µM. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 3) and statistical analysis [Student t-test (two-tailed) (***p < 0.001) or (*p < 0.05)]. (B) Bacteria were grown for 10 hours in BHI in the absence or presence of roseoflavin at a concentration of 1, 10 or 100 µM, respectively. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 3) and statistical analysis [Student t-test (two-tailed) (***p < 0.001)]. (C) L. monocytogenes was grown in BHI to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to Rli96 and tmRNA (control), respectively.](/cms/asset/43bd706b-ba5d-43f8-be36-08af941aaaf2/krnb_a_10915586_f0003.gif)
Figure 4 Mutations in the FMN riboswitch allows Listerial growth and expression of Imo1945 in the presence of roseoflavin. (A) Wild-type or M1 strains were grown for 10 hours in BHI in the absence or presence of roseoflavin at a concentration of 100 µM. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 6) and statistical analysis [Student t-test (two-tailed) (***p < 0.001)]. (B) L. monocytogenes wild-type or M1 strains were grown in BHI to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to Rli96, hly and tmRNA (control), respectively.
![Figure 4 Mutations in the FMN riboswitch allows Listerial growth and expression of Imo1945 in the presence of roseoflavin. (A) Wild-type or M1 strains were grown for 10 hours in BHI in the absence or presence of roseoflavin at a concentration of 100 µM. Growth was measured by optical density (OD600). Samples were compared to the control without added compound and shown as mean values with standard deviations (n = 6) and statistical analysis [Student t-test (two-tailed) (***p < 0.001)]. (B) L. monocytogenes wild-type or M1 strains were grown in BHI to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to Rli96, hly and tmRNA (control), respectively.](/cms/asset/7a0bc2ac-b87d-4322-a2e7-6d250617c59a/krnb_a_10915586_f0004.gif)
Figure 5 Treatment with roseoflavin results in a decreased expression of the virulence genes hly and actA in minimal medium. L. monocytogenes was grown in minimal medium to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to hly, actA and tmRNA (control), respectively.
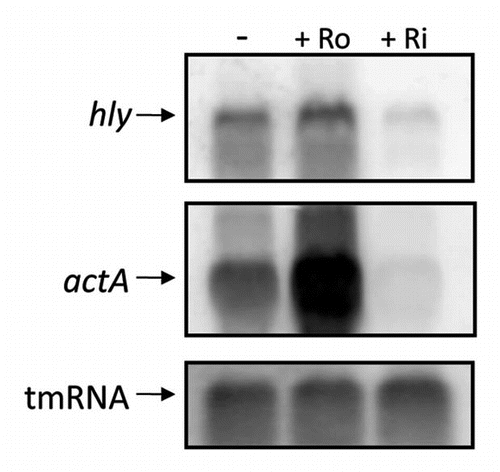
Figure 6 Roseoflavin does not decrease prfA monocistronic expression. (A) Schematic drawing of the prfA locus. Transcription of prfA initiates at two promoters, generating short prfA transcripts (hatched short lines). Active PrfA protein (solid spheres) can bind to the plcA promoter and stimulate transcription that generates plcA-prfA bi-cistronic transcript (long hatched line). (B) L. monocytogenes was grown in minimal medium to an OD600 = 0.25 when roseoflavin (Ro) or riboflavin (Ri) (100 µM) was added for ∼1.5 generations before RNA extraction. Northern blot was hybridized with PCR-generated, radioactively labeled DNA probes complementary to prfA and tmRNA (control), respectively.
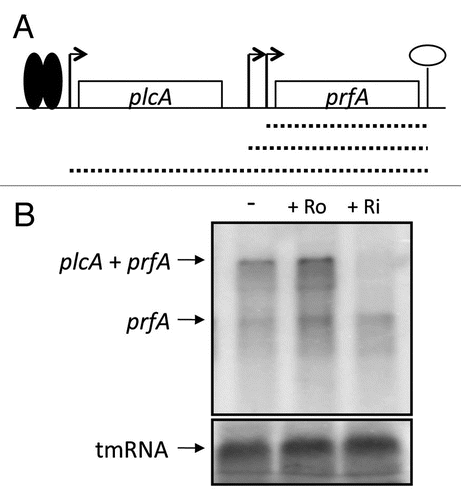
Figure 7 Riboflavin reduces the ability of L. monocytogenes to infect and proliferate inside HeLa cells. HeLa cells were infected with L. monocytogenes pretreated with roseoflavin or riboflavin (100 µM) for 1 generation at OD600 = 0.2. By viable counting, the amount of live bacteria was measured 1 hour or 4 hour post-infection, respectively, and these quantities were related to the number of live bacteria added to the HeLa cells and shown as mean values with standard deviations (n = 2). The invasivity of the treated bacteria were compared to untreated bacteria using Student t-test (two-tailed) (*p < 0.05).
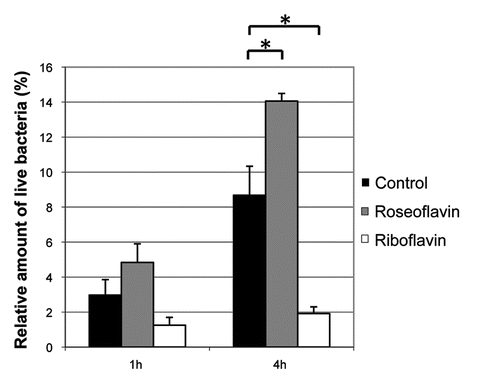
TABLE 1 Primers used in this study