Abstract
Epithelial tissues form a selective barrier that separates the external environment from the internal tissue milieu. Single epithelial cells are densely packed and associate via distinct intercellular junctions. Intercellular junction proteins not only control barrier properties of the epithelium but also play an important role in regulating epithelial homeostasis that encompasses cell proliferation, migration, differentiation and regulated shedding. Recent studies have revealed that several proteases target epithelial junction proteins during physiological maturation as well as in pathologic states such as inflammation and cancer. This review discusses mechanisms and biological consequences of transmembrane junction protein cleavage. The influence of junction protein cleavage products on pathogenesis of inflammation and cancer is discussed.
Introduction
Epithelial tissues form a physical barrier that separates the external environment and tissue compartments thereby playing a pivotal role in host defense. In simple epithelia such as in the gut, this barrier is formed by a single layer of cells joined by a series of intercellular junctions: tight junctions (TJs), adherens junctions (AJs) and desmosomes (DMs).Citation1 Intercellular junctions affiliate with the cytoskeleton that in turn controls junction function. It is now evident that intercellular junctions are dynamic structures that not only control the epithelial barrier, but also influence overall epithelial homeostasis.
TJs, the most apical epithelial junctions, are visualized as belt-like structures in a region where membranes from adjoining cells comes into very close apposition forming a selective seal that regulates paracellular movement of ions and solutes.Citation1,Citation2 In addition to its role as a gate, TJs also function as a fence that prevents free movement of proteins from the apical to the basolateral membrane. Thus, TJs contribute to maintenance of epithelial apical-basolateral polarity.Citation2 A vast repertoire of TJ proteins have been identified that can be broadly grouped into transmembrane, cytoplasmic plaque and signaling proteins. Key transmembrane proteins include claudins, occludin/tricellulin and the cortical thymocyte marker in Xenopus (CTX) family members (junctional adhesion molecule A (JAM-A), coxsackie adenovirus receptor (CAR) and CAR-like membrane protein (CLMP)). These transmembrane proteins affiliate with the underlying apical perijunctional F-actin belt via peripheral membrane proteins, such as zonula occludens (ZO)-1 and membrane-associated guanylate kinase with inverted orientation (MAGI)-1.Citation2-Citation6
AJs are localized immediately subjacent to the TJ and by electron microscopy appear as regions where membranes of two neighboring cells run parallel to each other.Citation1 Analogous to TJs, AJs circumscribe the cell periphery and play an important role regulating cell-cell adhesion. AJs consist of a central core of transmembrane proteins that include nectin, classical cadherin family members such as epithelial (E)-cadherin, neural (N)-cadherin, placental (P)-cadherin and cytoplasmic plaque proteins, which are linked to the actin cytoskeleton via plaque proteins.Citation7,Citation8 Plaque proteins in the AJ include afadin/AF-6 (for nectin), α-catenin and several members of the armadillo family that include β-catenin and p120-catenin (for classical cadherin).Citation8 It is noteworthy to mention that the AJ plaque protein β-catenin is one of the principal transcription factors that regulate epithelial proliferation and differentiation.Citation9,Citation10 Thus, establishment and maintenance of functional AJs is a central event that controls epithelial homeostasis.
DMs are spot-like adhesions located in the lateral membrane of the epithelial cellsCitation11,Citation12 and comprised of desmosomal cadherins: desmoglein (Dsg) and desmocollin (Dsc). To date, four isoforms of Dsg (Dsg1–4) and three isoforms of Dsc (Dsc1–3) have been identified in the human epidermis.Citation13 However, simple epithelia such as intestinal epithelium expresses only Dsg2 and Dsc2.Citation12,Citation14 Desmosomal cadherins indirectly associate with keratin intermediate filaments via plaque proteins, that include plakoglobin, plakophilin and the plakin family member, desmoplakin.Citation11 DMs provide resistance to mechanical stress in the epithelium and also in several nonepithelial tissues such as cardiac muscle and meninges.Citation12 In concert with other intercellular junctions, DMs are reservoirs for signaling molecules that control several biological processes.Citation11,Citation14,Citation15 Dysfunction of desmosomal cadherins has been linked to pathogenesis of diseases such as pemphigus vulgaris.Citation16-Citation18
Given their essential role in maintenance of the epithelial barrier, cell junctions need to function as dynamic structures that are continuously assembled and disassembled. The dynamic nature of cell junctions allows epithelial cells to respond to external stimuli by modifying strength and distribution of cell junction proteins. Remodeling of cell junctions can be achieved by mechanisms that include: de novo synthesis,Citation19 internalization,Citation20,Citation21 exocytosisCitation22 and proteolytic processing of structural proteins at cell junctions.Citation14,Citation23,Citation24 Indeed, proteolysis of transmembrane junction proteins by cellular and bacterial proteases has emerged as a novel mechanism to regulate intercellular junctions and epithelial homeostasis. In this review, we will focus on the mechanisms of transmembrane junction protein cleavage and their physiological, pathological and biological significance.
Extracellular Cleavage of Epithelial Transmembrane Junction Proteins
Proteases that mediate extracellular cleavage of transmembrane junction proteins
In epithelial tissues, the extracellular domains of the transmembrane junction proteins are located along the lateral membrane in the paracellular space formed by between two adjacent cells. In the intercellular space, ectodomains of two neighboring cells form cell junctions. Thus, the ectodomains are exposed to several insults including extracellular proteases referred to as “sheddases.” Sheddases can cleave unprotected extracellular domains of the proteins in the paracellular space (). Such proteases include diverse proteins derived from the epithelium, endothelium, immune cells and bacteria. Many sheddases are misregulated in pathological states such as inflammation and cancer. In fact, one of the mechanisms by which cancer cells overcome cell adhesion to induce cell proliferation and metastasis is by upregulating the expression/activity of several sheddases that are capable of cleaving the extracellular domain of cell adhesion molecules.Citation25,Citation26 As shown in , cleavage of transmembrane proteins most often occurs at defined sites close to transmembrane regions of junction proteins. Interestingly, the extracellar cleavage products can modulate epithelial cellular processes such as cell-cell adhesion, cell migration and cell proliferation. In this review, we discuss proteases that generate cleavage products of junctional proteins which exert biological effects on epithelial homeostasis. Matrix metalloprotease (MMP), a disintegrin and metalloprotease (ADAM), γ-secretase, serine proteases (kallikrein and plasmin) and bacterial proteases represent proteases that are involved in the cleavage of extracellular domain of junction proteins ( and ). It is noteworthy to mention that γ-secretase also functions as an intracellular protease for transmembrane junction proteins.Citation27
Figure 1. E-cadherin cleavage and its influence on epithelial homeostasis. This figure summarizes extracellular and intracellular cleavage of E-cadherin as detailed in the review.
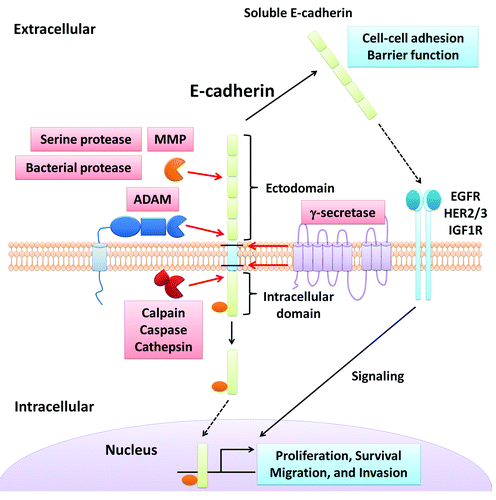
Figure 2. Potential cleavage sites in transmembrane junctional proteins. These figures demonstrate potential cleavage sites of representative transmembrane junctional proteins in epithelial cells. (A) Tight junction proteins; (B) E-cadherin; (C) desmoglein
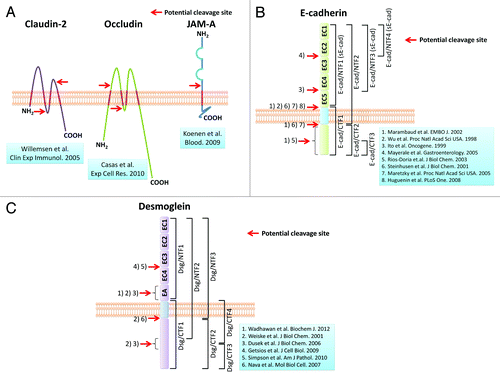
Table 1. Extracellular cleavage of transmembrane junction proteins in epithelial cells
Tight junction
Given the role of TJs as gatekeepers, cleavage of transmembrane proteins that form these structures is often associated with epithelial barrier breakdown. However, despite their importance in controlling epithelial barrier function, extracellular cleavage of integral proteins in TJs during physiological and pathological conditions is not well characterized. Huet et al. have reported that the induction of MMP9 by the extracellular matrix metalloprotease inducer (EMMPRIN; also termed CD147) results in cleavage of occludin extracellular domain and disruption of barrier function associated with pathogenesis of dry eye disease.Citation28 Willemsen et al. have also shown that interferon-gamma (IFNγ) activates cellular serine proteases that cleave claudin-2 extracellular domain.Citation29 Occludin has also been shown to be cleaved by several bacterial proteases including the cysteine protease Der p 1 from house dust mite,Citation30 haemagglutinin/protease (metalloprotease) from Vibrio Cholerae,Citation31 and secreted pore-forming toxin aerolysin produced by Aeromonas hydrophila.Citation32 Paracellular barrier compromise by haemagglutinin/protease and aerolysin has been linked to clinical diarrhea. Epithelial barrier disruption by house dust mite and pollen extract contributes to allergic diseases such as asthma.Citation30,Citation33,Citation34 Koenen et al. reported that cleavage and shedding of the TJ protein, JAM-A is mediated by ADAM17 and 10 during inflammation resulting in inhibition of neutrophil transendothelial diapedesis.Citation35 These findings highlight cleavage of transmembrane TJ proteins is essential mechanism that mediates disruption of epithelial barriers.
Adherens junction
The AJ intercellular space (~200 Å)Citation1 contains extracellular domains of classical-cadherin family members and nectins, which constitute the two major family of integral membrane proteins in this region. Nectins and cadherins control several physiological processes including cell-cell adhesion, cell signaling, proliferation and differentiation.Citation36-Citation38 For example, soluble E-cadherin (sE-cadherin) fragment resulting from cleavage of the E-cadherin extracellular domain by MMPsCitation39-Citation41 can function as a paracrine/autocrine signal to prevent cell death by activating epidermal growth factor receptor (EGFR) signaling and inhibiting apoptosis.Citation42 Shedding of sE-cadherin fragment has also been reported for other cellular proteases such as ADAMsCitation43 and γ-secretase.Citation27 In addition to cellular proteases, the extracellular domain of E-cadherin is targeted by several bacterial proteases that include HtrA protease from Helicobacter pylori,Citation44 fragilysin from Bacteroides fragilisCitation45 and the gingipains from Porphyromonas gingivali.Citation46 These bacterial proteases influence epithelial homeostasis analogous to cellular proteases. In addition to E-cadherin, other integral membrane AJ proteins such as the nectin-1 ectodomain are cleaved in neurons by MMPs.Citation47 The entire ectodomain of nectin-4 and nectin-1α are shed by MMPs in cancer and during cell spreading.Citation48,Citation49 The stability of these fragments in different body fluids suggests a putative role of these ectodomains as mediators of diverse biological processes.
Desmosome
Analogous to E-cadherin, desmosomal cadherins including Dsg and Dsc family members undergo extracellular domain cleavage during inflammation and cancer.Citation18,Citation50,Citation51 Indeed, extracellular domain cleavage of desmosomal cadherins has been reported to influence epithelial homeostasis.Citation18,Citation51,Citation52 In keratinocytes, where desmosomal cadherins are the major adhesion molecules, cleavage of desmosomal cadherins contributes to pathogenesis of diseases. For instance, the staphylococcal exfoliative toxin, which is causative agent of bullous impetigo and staphylococcal scalded skin syndrome have been shown to specifically target the ectodomain of Dsg1.Citation53,Citation54 In Netherton syndrome, which is an autosomal skin disease characterized by skin inflammation and scaling, Dsg1 cleavage occurs as a consequence of kallikrein hyperactivation due to the inactivation of the serine protease inhibitor of kazal type 5 (SPINK5).Citation55,Citation56 Additionally, shedding of desmosomal cadherin ectodomains by cellular proteases has been reported during cancer progression and metastasis implying a role of these molecules in modulating invasion and metastasis. For example, cleavage of Dsg2 in intestinal epithelial cells by matriptase has been associated with decreased cancer cell-cell adhesion.Citation57 ADAM17 and 10 have also been implicated in the shedding of Dsg2 during cancer development.Citation58 A similar mechanism has been described in the pathogenesis of oral squamous cell carcinoma where cleavage of Dsg1 by kallikrein 5 is necessary to reduce cell-cell adhesion in order to enhance cell migration.Citation51 Thus, understanding the mechanisms that regulate desmosomal cadherin cleavage is important in understanding the pathogenesis of several diseases.
Biological effects of epithelial junction protein extracellular cleavage fragments
Recent evidence suggests that cleavage fragment of junctional proteins possess different biological activities.Citation26 These fragment not only control normal homeostatic event in the epithelia such as cell proliferation, migration and apoptosis but also contribute to pathogenesis of mucosal inflammation and cancer. In this section, we describe the biological effects of soluble intercellular junction protein cleavage fragments.
Cell-cell adhesion
Cell-cell adhesion in epithelial cells is mediated by AJ and desmosomal proteins. It’s well known that disruption of cell-cell adhesion contributes to metastasis of cancer cells. Several reports have shown that extracellular cleavage of intercellular junction proteins results in loss of cell-cell adhesion.Citation59,Citation60 Moreover, recent findings have also shown that soluble extracellular cleavage products of intercellular junction proteins can regulate epithelial cell-cell adhesion by paracrine or autocrine signaling. Wheelock et al. reported that the 80 kd sE-cadherin fragment disrupts cell-cell adhesion in a human mammary carcinoma cell line.Citation61 Symowicz et al. have also demonstrated that recombinant sE-cadherin promotes cell junction disruption and metastasis of ovarian carcinoma cells.Citation41 Additionally, extracellular E-cadherin fragments released by serine proteases, plasmin and kallikrein 7 inhibit cellular aggregation.Citation62,Citation63 These findings suggest that extracellular E-cadherin fragments have functional effects that inhibit endogenous cell-cell junction protein function. However, it still remains unknown how these cleavage products interact with cell-cell junction proteins and whether or not extracellular cleavage products of other intercellular junction proteins such as nectins or desmosomal cadherins have similar biological effects on cell-cell adhesion.
Barrier function
Epithelial tissues provide physical and permeability barriers in vertebrates. In simple epithelium the barrier is formed by epithelial cells where the paracellular space is sealed by intercellular contacts. MMPs actively participate in the disruption of epithelial barrier under several conditions. For example, during inflammation, proinflammatory cytokines released in the milieu of ocular epithelium enhance MMP9 expression and activation that promotes cleavage of the TJ protein occludin thereby resulting in increased corneal epithelial permeability.Citation28,Citation64-Citation66 Of interest MMP9 expression and activity has been directly linked to dry eyes in human patients with ocular rosacea and rheumatoid arthritis.Citation64,Citation65 Moreover, virulence factors can also perturb epithelial barrier function by inducing cleavage of specific transmembrane epithelial junction proteins. The effects of these bacterial proteases are broad in range and some are very well characterized. Fragilysin from Bacteroides fragilis cleaves the extracellular domain of E-cadherin to disrupt barrier function.Citation45 In contrast, the zinc-containing metalloprotease from Vibrio cholerae decreases epithelial barrier function by targeting the extracellular domain of occludin.Citation31 In addition to its role in compromising the epithelial barrier, mimetic peptides of transmembrane junction proteins and virus-derived proteins can transiently modulate epithelial barrier function. Interestingly, the efficiency and reliability of this system has been used in recent years to transiently compromise the paracellular pathway in order to promote drug delivery across the epithelium.Citation67-Citation71
Migration and invasion
Epithelial cells are less mobile than most other cell types. Disruption of cell adhesion to enhance cell migration and invasion is an integral event in cancer metastasis.Citation72 As mentioned above, cleavage of transmembrane junction proteins by proteases reduces cell-cell adhesion and directly enhances cell migration and invasion.Citation59,Citation60 Interestingly, it has been shown that the extracellular cleavage products from intercellular junction proteins can also induce cell migration and invasion. The role of sE-cadherin in cell migration and invasion is well known.Citation40,Citation62,Citation73,Citation74 Most recently, Brouxhon et al. have demonstrated that sE-cadherin promotes cell migration and tumor invasion by activating MMP2 and 9 in murine skin squamous carcinoma cell lines.Citation75 Furthermore, in human lung tumor cells, the presence of sE-cadherin in conditioned media and E-cadherin HAV peptides simulating E-cadherin EC1 domain induce MMP2, 9 and MT1-MMP transcription and activation resulting in cell invasion.Citation76 Taken together, these findings suggest that sE-cadherin has functional properties that promote cell migration and invasion by inducing MMP activity. Increased junction protein cleavage, reported in inflammation, exerts biological effects on mucosal homeostasis. Soluble JAM-A ectodomain fragments have been reported to modulate endothelial cell migration and prevent neutrophil extravasation.Citation35 However, mechanisms by which soluble extracellular junction protein cleavage fragments modulate cell migration are complex and not well understood.
Cell proliferation and apoptosis
Several reports have linked junction extracellular cleavage products to cell proliferation and apoptosis.Citation42,Citation59,Citation75,Citation77 These studies have also highlighted underlying signaling pathways that mediate the biological effect of junction protein cleavage fragments. In skin cancer cells, exogenous sE-cadherin can interact with EGFR and human epidermal growth factor receptor (HER) 2 to activate them and their downstream singnaling proteins: phosphoinositide-3 kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) and mitogen-activated protein kinase (MAPK). Ultimately, the cleavage fragments induce cell proliferation.Citation75 Najy et al. have also shown the interaction between exogenous sE-cadherin and HER2-HER3 heterodimer in breast cancer cells where sE-cadherin promotes cell proliferation via HER2-HER3 and extracellular signal-regulated kinase (ERK) activation.Citation77 In addition to proliferation, recent evidence indicates that extracellular cleavage products can regulate cell survival. For example, Inge et al. have revealed that sE-cadherin can suppress apoptosis thereby promote cell survival and these effects are mediated by activation of EGFR signaling via PI3K/Akt and ERK1/2.Citation42 However, cleavage or loss of the transmembrane proteins in cell junctions has also been linked to the presence or appearance of pro-apoptotic signals.Citation78 In fact, metalloprotease cleavage of desmosomal cadherins during keratinocyte apoptosis has been reported.Citation79 Additionally, and in simple epithelium, cleavage of E-cadherin by sheddasses is an essential step in the onset of apoptosis.Citation80,Citation81
Intracellular Cleavage of Epithelial Transmembrane Junction Proteins
Proteases that mediate intracellular cleavage of junction proteins
Transmembrane junction proteins associate with an underlying actin cytoskeleton via scaffold proteins that constrains their intracellular domains. Intracellular domains can also be targeted by proteases that are in the vicinity of the cytoplasmic surface of the plasmamembrane.
Several proteases that are activated during inflammation and cancer mediate intracellular cleavage of cell-cell junction proteins. Additionally, increased expression/activity of proteases that cleave adhesion proteins facilitates cancer cell proliferation and metastasis.Citation25,Citation26 Interestingly, following extracellular domain shedding, the membrane-associated proteins undergo proteolytic processing thereby generating soluble intracellular fragments that exert biological effects. Cleavage of such protein fragments is mediated by intracellular and membrane associated proteases such as calpain, caspase, cathepsin and γ-secretase.Citation27 Intracellular proteases promote intracellular cleavage of junction proteins that in turn induces cytoskeletal restructuring as well as modification in scaffold and signaling proteins to maintain epithelial homeostasis (). Recent reports have highlighted biological effects of intracellular cleavage fragments of junction proteins ().
Table 2. Intracellular cleavage of transmembrane junction protein in epithelial cells
Tight junction
Little is known about the intracellular cleavage of TJ proteins. Sumitomo et al. have demonstrated that calpain activated by Streptolysin S from Group A streptococcus, cleaves occludin and disrupts barrier function of intestinal epithelial cells and keratinocytes.Citation82 Willemsen et al. have also shown that cellular serine protease activated by IFN-γ treatment can cleave intracellular domain of claudin-2.Citation29 Additionally, toll-like receptor (TLR) 2-induced calpain cleaves intracellular domain of occludin and promotes polymorphonuclear leukocyte transmigration.Citation83
Adherens junction
The intracellular fragment of E-cadherin that remains attached to the cellular membrane after cleavage by membrane-bound metalloproteases, generates a cytosolic fragment (33 kDa) when γ-secretase is activated. The 33 kDa cytosolic fragment promotes intracellular signaling and cell proliferation.Citation27,Citation84 In addition to cleavage by membrane associated proteases, the cytosolic domain of E-cadherin has been shown to be further cleaved by intracellular proteases such as calpain.Citation23 However, the fragments generated by calpain have been shown to induce apoptosis instead of proliferation in epithelial cells.Citation81 Interestingly, it has also been proposed that during apoptosis the 33 kDa fragment generated by γ-secretase is also targeted by caspase-3 to generate a 29 kDa fragment. However, the function of this cleavage fragment has not been characterized, but its localization and distribution suggest a putative role in the regulation of epithelial cell apoptosis.Citation80 Thus, proteolysis of epithelial cadherin allows the formation of several intracellular fragments that have biological activities. Furthermore, E-cadherin intracellular fragments are important in activating signaling pathways that control physiological and pathological events.
Desmosome
In simple epithelium, cleavage of desmosomal cadherins by cysteine proteases has been shown to be an important step in the disassembly of the DMs.Citation85 Desmosomal disassembly may directly contribute to the detachment of the dead cells observed in epithelial tissues. The cytosolic fragments generated by Dsg2 cleavage have been observed in human colon tissues.Citation86 We have observed that an intracellular Dsg2 cleavage fragment promotes apoptosis in the intestinal epithelium.Citation14 We have also reported a similar Dsg2 cleavage fragment in nasal polyps after cytokine exposure.Citation87 However, in addition to apoptosis, we cannot rule out the possibility that the cytosolic fragments generated after Dsg2 cleavage serve to promote cell proliferation. In fact, Brennan et al. have demonstrated that Dsg2 proteolytic products are elevated in vivo in skin tumors from transgenic mice overexpressing Dsg2.Citation88 These results suggest that the cytosolic fragment of Dsg2 may be responsible for the regulation of several mitogenic signaling pathways such as Akt, MAPK and the signal transducer and activator of transcription (STAT) 3.Citation89
Biological effects of junction protein intracellular cleavage products
As described above, several intracellular proteases including γ-secretase, calpain and caspase are responsible for cleavage of intercellular junction proteins that in turn exert biological effects. We have previously observed that an intracellular Dsg2 cleavage fragment generated by serine proteases promotes apoptosis in intestinal epithelial cells.Citation14 Some cleavage fragments can translocate to the nucleus to regulate transcriptional activity and cell survival. For instance, 35 kDa E-cadherin C-terminal fragment (CTF) generated by presenilin 1 regulates β-catenin/T cell factor (Tcf)-4 transcriptional activity.Citation90 E-cadherin/CTF translocates to the nucleus to influence Kaiso-regulated gene transcription. Nuclear localization of E-cadherin/CTF is also enhanced by p120-catenin.Citation91 Furthermore, presenilin 1 induces E-cadherin cytoplasmic cleavage, generating products that promote disassembly of E-cadherin-catenin complex by sequestering β-catenin in the cytosol.Citation27 Additionally, calpain induced cleavage of intracellular E-cadherin domain generates a 100 kDa cleavage product that regulates cell survival in prostate epithelial cells.Citation81 Similar to E-cadherin, presenillin promotes the release of the CTF in N-cadherin which also plays an important role in the regulation of β-catenin signaling.Citation92
In addition to cadherins, intracellular domains of epithelial cell adhesion molecule (EpCAM) generated by presenilin 2 translocates to the nucleus with four and a half LIM-domain protein (FHL) 2 and β-catenin to promote lymphoid enhancer-binding factor (Lef)-1 transactivation and epithelial proliferation.Citation93 Interestingly, truncated Dsg1 and 2 generated by extracellular domain cleavage can influence cell-surface desmosomal cadherins through the interaction of raft-associated protein, caveolin-1 and plakoglobin.Citation50,Citation88
Signaling Pathways Triggered by Cleavage of Transmembrane Junction Proteins
Transmembrane junction proteins regulate cell proliferation, differentiation and apoptosis by controlling several signaling pathways. The mechanism by which the cleavage fragments regulate these processes is incompletely understood. Dimerization and autophosphorylation of receptor tyrosine kinases (RTKs) by the extracellular cleavage products of cadherin family members can regulate epithelial homeostasis. For example, sE-cadherin results in RTK ligand-independent dimerization, activation of the receptor and stimulation of several intracellular signaling pathways including MAPK, PI3K, Akt and mTOR.Citation94,Citation95 These signaling pathways influence biological events such as cell proliferation,Citation94,Citation95 growth,Citation94 and cell migration.Citation96 As a consequence of RTK interaction with sE-cadherin, activation of pro-proliferative and anti-apoptotic signaling pathways was observed. Thus, the presence of sE-cadherin in the serum has been associated with poor prognosis in cancer patients.Citation77,Citation97 However, given that shedding of the cadherin ectodomain has also been reported in physiological conditions or in other pathologies such as inflammation,Citation50,Citation52-Citation54,Citation98-Citation100 it is easy to speculate that these soluble fragments have additional biological functions that need to be investigated.
Cleavage of transmembrane junctional proteins can directly trigger the activation of pro-proliferative signaling by releasing intracellular scaffolding molecules that often associate with their intracellular domains. For instance, intracellular cleavage of E-cadherin and N-cadherin promotes translocation of β-catenin from the plasmamembrane to nucleus thereby resulting in β-catenin/TCF transactivation and increase in cell proliferation.Citation43,Citation84,Citation92 In addition to its role in cell proliferation, β-catenin transactivation can also increase the expression and secretion of metalloproteases.Citation101,Citation102 Thus, it is tempting to speculate that β-catenin redistribution after E-cadherin or N-cadherin cleavage could result in the activation of metalloproteases that in turn enhance shedding of their ectodomains. This process will increase epithelial cell proliferation through stimulating RTK receptors in a paracrine manner.
Relevance to Pathologic States
Cancer
A continuous turnover of intercellular junction proteins in healthy individuals is required for the maintenance of epithelial tissues and therefore, a low level of E-cadherin cleavage product has been observed in the healthy serum.Citation99 However, in pathological conditions such as inflammation and cancer, proteolytic activity is increased.Citation103 Since cleavage products have biological activity that influences cell-cell adhesion, migration and proliferation, they contribute to the pathogenesis of disease. As a result, increased soluble junctional proteins such as E-cadherin,Citation40,Citation99,Citation104,Citation106-Citation116 N-cadherin,Citation105 and nectinCitation49 can be observed in body fluids (serum, urine and ascites) and cancer tissue of patients. Given the ectodomain stability of cell junction proteins in several body fluids, the detection of cleavage products as biomarkers for disease has been considered. For example, increased sE-cadherin is seen in serum or at other sites (cancer tissue, urine, cyst and ascites) in cancer patients. Such cleavage product has been detected in cancers from many areas that include bladder,Citation106,Citation107 colorectal,Citation108,Citation109 esophageal squamous cell carcinoma,Citation104 gastric,Citation99,Citation110 liver,Citation99,Citation111 lung,Citation112,Citation113 ovarian,Citation41,Citation114,Citation115 prostate,Citation116 and skin tissue.Citation117 Given the biological function of intercellular junction protein ectodomains, their upregulation may play a role in the progression of cancer and metastasis. However, some reports have failed to detect increased serum E-cadherin cleaved products in some cancer patients.Citation118-Citation120 Thus, additional studies are needed to verify the role of such cleavage products in cancer pathogenesis and detection. Analogous to E-cadherin, the ectodomains of nectin-4 are increased in the serum of patients with metastatic ductal breast carcinoma compared with healthy subjects.Citation49 Furthermore, soluble N-cadherin is significantly elevated in serum of prostate cancer patients.Citation105 These findings indicate the potential use of such products as diagnostic tools.
Inflammation
Knowledge of junctional protein cleavage fragments in inflammation is limited compared with cancer reports. Mayerle et al. have shown that leukocyte elastase cleaves extracellular domain of E-cadherin and disrupt cell-cell contacts in the rat pancreas, thereby promoting leukocyte transmigration into epithelial tissues in the initial phase of experimental pancreatitis.Citation100 Furthermore, extracellular cleavage of Dsg1 by Staphylococcus aureus exfoliative toxin A (serine protease) disrupts cell-cell adhesion, which leads to bullous impetigo or Staphylococcal scalded skin syndrome.Citation50,Citation52-Citation54 These reports suggest that the extracellular cleavage of intercellular junction proteins disrupts cell-cell adhesion contributing to loss of epithelial barrier function and mucosal inflammation. However, the utility of detecting junctional protein cleavage products as inflammation biomarkers remains incompletely understood. Pittard et al. identified increased concentration of sE-cadherin in patients with systemic inflammatory response syndrome and multi-organ dysfunction.Citation98 On the contrary, Weiss at al. did not observe significant difference in serum sE-cadherin in healthy control individuals and patients with inflammatory bowel disease.Citation109 The presence of the ectodomains of other junction proteins such as JAM-A and JAM-CCitation121 in body fluids suggests a putative role of these fragments as diagnostic tools in such pathologies.
Others
Cleavage of transmembrane junction proteins has been observed in several other pathologies. Dry eye disease is a common disease that develops as a result of changes in tear fluid, leading to osmotic stress and perturbed epithelial barrier function. Huet et al. have shown a relationship of MMP9-mediated cleavage of occludin in human corneal epithelial cells and dry eye disease.Citation28 In HIV infection, abnormal distribution of E-cadherin has been observed in the intestinal epithelium, which leads to increased systemic levels of sE-cadherin. Additionally, sE-cadherin level in plasma correlates with viral load in HIV patients. Interestingly, sE-cadherin can inhibit HIV-1-specific antiviral activity of CD8+ T-cell function.Citation122 Thus, cleavage products of intercellular junctional proteins also regulate immune response. Additionally, it has to be noted that some pathological states induced by bacterial pathogens such as diarrheaCitation45 and skin blister formationCitation123 are mediated by targeting extracellular domains of cell adhesion molecules by bacterial proteases.
Conclusion
A continuous turnover of transmembrane intercellular junction proteins is observed in healthy individuals and is required for the maintenance of epithelial barriers. Physiologically low levels of cleavage products have been identified in the serum. The cleavage products are generated by a variety of proteases during epithelial cell junction rearrangement. Extracellular proteases such as MMP, ADAM, γ-secretase, serine and bacterial proteases can cleave the ectodomain of the proteins. On the other hand, proteases including calpain and caspase cleave intracellular domain of junction transmembrane proteins. Interestingly, the cleavage products generated by these proteases have distinct biological functions. In pathological conditions such as cancer and inflammation, increased junction protein cleavage products are often detected in the tissue and body fluids. These cleavage products influence epithelial behavior that includes proliferation, migration and apoptosis. In addition to exerting biological activity, detection of junction protein cleavage fragment can serve as biomarkers to follow disease progression.
| Abbreviations: | ||
| TJ | = | tight junction |
| AJ | = | adherens junction |
| DM | = | desmosome |
| CTX | = | cortical thymocyte marker in Xenopus |
| JAM-A | = | junctional adhesion molecule A |
| CAR | = | coxsackie adenovirus receptor |
| CLMP | = | CAR-like membrane protein |
| ZO | = | zonula occludens |
| MAGI | = | membrane-associated guanylate kinase with inverted orientation |
| E-cadherin | = | epithelial-cadherin |
| N-cadherin | = | neural-cadherin |
| P-cadherin | = | placental-cadherin |
| AF | = | afadin |
| Dsg | = | desmoglein |
| Dsc | = | desmocollin |
| MMP | = | matrix metalloprotease |
| ADAM | = | a disintegrin and metalloprotease |
| EMMPRIN | = | extracellular matrix metalloprotease inducer |
| IFN-γ | = | interferon-gamma |
| sE-cadherin | = | soluble E-cadherin |
| EGFR | = | epidermal growth factor receptor |
| HtrA | = | high temperature requirement protease A |
| SPINK5 | = | serine protease inhibitor of kazal type 5 |
| HAV | = | His-Ala-Val |
| EC | = | extracellular |
| MT1-MMP | = | membrane-type matrix metalloprotease |
| HER | = | human epidermal growth factor receptor |
| PI3K | = | phosphoinositide-3 kinase |
| mTOR | = | mammalian target of rapamycin |
| MAPK | = | mitogen-activated protein kinase |
| ERK | = | extracellular signal-regulated kinase |
| TLR | = | toll-like receptor |
| STAT | = | signal transducer and activator of transcription |
| CTF | = | C-terminal fragment |
| Tcf | = | T cell factor |
| EpCAM | = | epithelial cell adhesion molecule |
| FHL2 | = | four and a half LIM-domain protein |
| Lef | = | lymphoid enhancer-binding factor |
| RTK | = | receptor tyrosine kinases |
| HIV | = | human immunodeficiency virus |
| TACE | = | tumor necrosis factor-α-converting enzyme |
| TGFβ | = | transforming growth factor-beta |
| IL-1β | = | interleukin-1 beta |
| LPS | = | lipopolysaccharide |
| TNFα | = | tumor necrosis factor-alpha |
| PMA | = | phorbol-12-myristate-13-acetate |
| PKD1 | = | protein kinase D1 |
| HGF/SF | = | hepatocyte growth factor/scatter factor |
| EGF | = | epidermal growth factor |
| LPA | = | lysophosphatidic acid |
| TPA | = | 12-O-tetradecanoylphorbol-13-acetate |
| PMN | = | polymorphonuclear leukocyte |
| IGF1R | = | insulin-like growth factor 1 receptor |
| NTF | = | N-terminal fragment |
| EA | = | extracellular anchor |
Acknowledgments
This work was supported by an American Gastroenterological Association Research Scholar Award (P.N.), a Crohn’s and Colitis Foundation of America Career Development Award (P.N.) and a CONACyT grant 175854 (P.N.), the 12th Glaxo-SmithKline International Award from Japan Society of Immunology and Allergology in Otolaryngology (R.K.) and National Institutes of Health grants DK059888 and DK055679 (A.N.).
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
References
- Farquhar MG, Palade GE. Junctional complexes in various epithelia. J Cell Biol 1963; 17:375 - 412; http://dx.doi.org/10.1083/jcb.17.2.375; PMID: 13944428
- Tsukita S, Furuse M, Itoh M. Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol 2001; 2:285 - 93; http://dx.doi.org/10.1038/35067088; PMID: 11283726
- Sawada N, Murata M, Kikuchi K, Osanai M, Tobioka H, Kojima T, et al. Tight junctions and human diseases. Med Electron Microsc 2003; 36:147 - 56; http://dx.doi.org/10.1007/s00795-003-0219-y; PMID: 14505058
- Schneeberger EE, Lynch RD. The tight junction: a multifunctional complex. Am J Physiol Cell Physiol 2004; 286:C1213 - 28; http://dx.doi.org/10.1152/ajpcell.00558.2003; PMID: 15151915
- Ikenouchi J, Furuse M, Furuse K, Sasaki H, Tsukita S, Tsukita S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J Cell Biol 2005; 171:939 - 45; http://dx.doi.org/10.1083/jcb.200510043; PMID: 16365161
- Balda MS, Matter K. Tight junctions at a glance. J Cell Sci 2008; 121:3677 - 82; http://dx.doi.org/10.1242/jcs.023887; PMID: 18987354
- Niessen CM. Tight junctions/adherens junctions: basic structure and function. J Invest Dermatol 2007; 127:2525 - 32; http://dx.doi.org/10.1038/sj.jid.5700865; PMID: 17934504
- Harris TJ, Tepass U. Adherens junctions: from molecules to morphogenesis. Nat Rev Mol Cell Biol 2010; 11:502 - 14; http://dx.doi.org/10.1038/nrm2927; PMID: 20571587
- Nelson WJ, Nusse R. Convergence of Wnt, β-catenin, and cadherin pathways. Science 2004; 303:1483 - 7; http://dx.doi.org/10.1126/science.1094291; PMID: 15001769
- Brembeck FH, Rosário M, Birchmeier W. Balancing cell adhesion and Wnt signaling, the key role of β-catenin. Curr Opin Genet Dev 2006; 16:51 - 9; http://dx.doi.org/10.1016/j.gde.2005.12.007; PMID: 16377174
- Dusek RL, Godsel LM, Green KJ. Discriminating roles of desmosomal cadherins: beyond desmosomal adhesion. J Dermatol Sci 2007; 45:7 - 21; http://dx.doi.org/10.1016/j.jdermsci.2006.10.006; PMID: 17141479
- Holthöfer B, Windoffer R, Troyanovsky S, Leube RE. Structure and function of desmosomes. Int Rev Cytol 2007; 264:65 - 163; http://dx.doi.org/10.1016/S0074-7696(07)64003-0; PMID: 17964922
- Green KJ, Simpson CL. Desmosomes: new perspectives on a classic. J Invest Dermatol 2007; 127:2499 - 515; http://dx.doi.org/10.1038/sj.jid.5701015; PMID: 17934502
- Nava P, Laukoetter MG, Hopkins AM, Laur O, Gerner-Smidt K, Green KJ, et al. Desmoglein-2: a novel regulator of apoptosis in the intestinal epithelium. Mol Biol Cell 2007; 18:4565 - 78; http://dx.doi.org/10.1091/mbc.E07-05-0426; PMID: 17804817
- Jamora C, Fuchs E. Intercellular adhesion, signalling and the cytoskeleton. Nat Cell Biol 2002; 4:E101 - 8; http://dx.doi.org/10.1038/ncb0402-e101; PMID: 11944044
- Amagai M, Klaus-Kovtun V, Stanley JR. Autoantibodies against a novel epithelial cadherin in pemphigus vulgaris, a disease of cell adhesion. Cell 1991; 67:869 - 77; http://dx.doi.org/10.1016/0092-8674(91)90360-B; PMID: 1720352
- Allen EM, Giudice GJ, Diaz LA. Subclass reactivity of pemphigus foliaceus autoantibodies with recombinant human desmoglein. J Invest Dermatol 1993; 100:685 - 91; http://dx.doi.org/10.1111/1523-1747.ep12472348; PMID: 8491991
- Waschke J. The desmosome and pemphigus. Histochem Cell Biol 2008; 130:21 - 54; http://dx.doi.org/10.1007/s00418-008-0420-0; PMID: 18386043
- McCarthy KM, Francis SA, McCormack JM, Lai J, Rogers RA, Skare IB, et al. Inducible expression of claudin-1-myc but not occludin-VSV-G results in aberrant tight junction strand formation in MDCK cells. J Cell Sci 2000; 113:3387 - 98; PMID: 10984430
- Ivanov AI, Nusrat A, Parkos CA. Endocytosis of the apical junctional complex: mechanisms and possible roles in regulation of epithelial barriers. Bioessays 2005; 27:356 - 65; http://dx.doi.org/10.1002/bies.20203; PMID: 15770686
- Utech M, Ivanov AI, Samarin SN, Bruewer M, Turner JR, Mrsny RJ, et al. Mechanism of IFN-γ-induced endocytosis of tight junction proteins: myosin II-dependent vacuolarization of the apical plasma membrane. Mol Biol Cell 2005; 16:5040 - 52; http://dx.doi.org/10.1091/mbc.E05-03-0193; PMID: 16055505
- Andersen NJ, Yeaman C. Sec3-containing exocyst complex is required for desmosome assembly in mammalian epithelial cells. Mol Biol Cell 2010; 21:152 - 64; http://dx.doi.org/10.1091/mbc.E09-06-0459; PMID: 19889837
- Rios-Doria J, Day KC, Kuefer R, Rashid MG, Chinnaiyan AM, Rubin MA, et al. The role of calpain in the proteolytic cleavage of E-cadherin in prostate and mammary epithelial cells. J Biol Chem 2003; 278:1372 - 9; http://dx.doi.org/10.1074/jbc.M208772200; PMID: 12393869
- Kowalczyk AP, Nanes BA. Adherens junction turnover: regulating adhesion through cadherin endocytosis, degradation, and recycling. Subcell Biochem 2012; 60:197 - 222; http://dx.doi.org/10.1007/978-94-007-4186-7_9; PMID: 22674073
- McCawley LJ, Matrisian LM. Matrix metalloproteinases: they’re not just for matrix anymore!. Curr Opin Cell Biol 2001; 13:534 - 40; http://dx.doi.org/10.1016/S0955-0674(00)00248-9; PMID: 11544020
- Grabowska MM, Day ML. Soluble E-cadherin: more than a symptom of disease. Front Biosci 2012; 17:1948 - 64; http://dx.doi.org/10.2741/4031; PMID: 22201848
- Marambaud P, Shioi J, Serban G, Georgakopoulos A, Sarner S, Nagy V, et al. A presenilin-1/γ-secretase cleavage releases the E-cadherin intracellular domain and regulates disassembly of adherens junctions. EMBO J 2002; 21:1948 - 56; http://dx.doi.org/10.1093/emboj/21.8.1948; PMID: 11953314
- Huet E, Vallée B, Delbé J, Mourah S, Prulière-Escabasse V, Tremouilleres M, et al. EMMPRIN modulates epithelial barrier function through a MMP-mediated occludin cleavage: implications in dry eye disease. Am J Pathol 2011; 179:1278 - 86; http://dx.doi.org/10.1016/j.ajpath.2011.05.036; PMID: 21777561
- Willemsen LE, Hoetjes JP, van Deventer SJ, van Tol EA. Abrogation of IFN-γ mediated epithelial barrier disruption by serine protease inhibition. Clin Exp Immunol 2005; 142:275 - 84; http://dx.doi.org/10.1111/j.1365-2249.2005.02906.x; PMID: 16232214
- Wan H, Winton HL, Soeller C, Gruenert DC, Thompson PJ, Cannell MB, et al. Quantitative structural and biochemical analyses of tight junction dynamics following exposure of epithelial cells to house dust mite allergen Der p 1. Clin Exp Allergy 2000; 30:685 - 98; http://dx.doi.org/10.1046/j.1365-2222.2000.00820.x; PMID: 10792361
- Wu Z, Nybom P, Magnusson KE. Distinct effects of Vibrio cholerae haemagglutinin/protease on the structure and localization of the tight junction-associated proteins occludin and ZO-1. Cell Microbiol 2000; 2:11 - 7; http://dx.doi.org/10.1046/j.1462-5822.2000.00025.x; PMID: 11207559
- Bücker R, Krug SM, Rosenthal R, Günzel D, Fromm A, Zeitz M, et al. Aerolysin from Aeromonas hydrophila perturbs tight junction integrity and cell lesion repair in intestinal epithelial HT-29/B6 cells. J Infect Dis 2011; 204:1283 - 92; http://dx.doi.org/10.1093/infdis/jir504; PMID: 21917902
- Runswick S, Mitchell T, Davies P, Robinson C, Garrod DR. Pollen proteolytic enzymes degrade tight junctions. Respirology 2007; 12:834 - 42; http://dx.doi.org/10.1111/j.1440-1843.2007.01175.x; PMID: 17986111
- Heijink IH, van Oosterhout A, Kapus A. Epidermal growth factor receptor signalling contributes to house dust mite-induced epithelial barrier dysfunction. Eur Respir J 2010; 36:1016 - 26; http://dx.doi.org/10.1183/09031936.00125809; PMID: 20351035
- Koenen RR, Pruessmeyer J, Soehnlein O, Fraemohs L, Zernecke A, Schwarz N, et al. Regulated release and functional modulation of junctional adhesion molecule A by disintegrin metalloproteinases. Blood 2009; 113:4799 - 809; http://dx.doi.org/10.1182/blood-2008-04-152330; PMID: 19258599
- Takai Y, Irie K, Shimizu K, Sakisaka T, Ikeda W. Nectins and nectin-like molecules: roles in cell adhesion, migration, and polarization. Cancer Sci 2003; 94:655 - 67; http://dx.doi.org/10.1111/j.1349-7006.2003.tb01499.x; PMID: 12901789
- Saito M, Tucker DK, Kohlhorst D, Niessen CM, Kowalczyk AP. Classical and desmosomal cadherins at a glance. J Cell Sci 2012; 125:2547 - 52; http://dx.doi.org/10.1242/jcs.066654; PMID: 22833291
- Le Bras GF, Taubenslag KJ, Andl CD. The regulation of cell-cell adhesion during epithelial-mesenchymal transition, motility and tumor progression. Cell Adh Migr 2012; 6:365 - 73; http://dx.doi.org/10.4161/cam.21326; PMID: 22796940
- Davies G, Jiang WG, Mason MD. Matrilysin mediates extracellular cleavage of E-cadherin from prostate cancer cells: a key mechanism in hepatocyte growth factor/scatter factor-induced cell-cell dissociation and in vitro invasion. Clin Cancer Res 2001; 7:3289 - 97; PMID: 11595727
- Noë V, Fingleton B, Jacobs K, Crawford HC, Vermeulen S, Steelant W, et al. Release of an invasion promoter E-cadherin fragment by matrilysin and stromelysin-1. J Cell Sci 2001; 114:111 - 8; PMID: 11112695
- Symowicz J, Adley BP, Gleason KJ, Johnson JJ, Ghosh S, Fishman DA, et al. Engagement of collagen-binding integrins promotes matrix metalloproteinase-9-dependent E-cadherin ectodomain shedding in ovarian carcinoma cells. Cancer Res 2007; 67:2030 - 9; http://dx.doi.org/10.1158/0008-5472.CAN-06-2808; PMID: 17332331
- Inge LJ, Barwe SP, D’Ambrosio J, Gopal J, Lu K, Ryazantsev S, et al. Soluble E-cadherin promotes cell survival by activating epidermal growth factor receptor. Exp Cell Res 2011; 317:838 - 48; http://dx.doi.org/10.1016/j.yexcr.2010.12.025; PMID: 21211535
- Maretzky T, Reiss K, Ludwig A, Buchholz J, Scholz F, Proksch E, et al. ADAM10 mediates E-cadherin shedding and regulates epithelial cell-cell adhesion, migration, and β-catenin translocation. Proc Natl Acad Sci U S A 2005; 102:9182 - 7; http://dx.doi.org/10.1073/pnas.0500918102; PMID: 15958533
- Hoy B, Geppert T, Boehm M, Reisen F, Plattner P, Gadermaier G, et al. Distinct roles of secreted HtrA proteases from gram-negative pathogens in cleaving the junctional protein and tumor suppressor E-cadherin. J Biol Chem 2012; 287:10115 - 20; http://dx.doi.org/10.1074/jbc.C111.333419; PMID: 22337879
- Wu S, Lim KC, Huang J, Saidi RF, Sears CL. Bacteroides fragilis enterotoxin cleaves the zonula adherens protein, E-cadherin. Proc Natl Acad Sci U S A 1998; 95:14979 - 84; http://dx.doi.org/10.1073/pnas.95.25.14979; PMID: 9844001
- Katz J, Yang QB, Zhang P, Potempa J, Travis J, Michalek SM, et al. Hydrolysis of epithelial junctional proteins by Porphyromonas gingivalis gingipains. Infect Immun 2002; 70:2512 - 8; http://dx.doi.org/10.1128/IAI.70.5.2512-2518.2002; PMID: 11953390
- Lim ST, Chang A, Giuliano RE, Federoff HJ. Ectodomain shedding of nectin-1 regulates the maintenance of dendritic spine density. J Neurochem 2012; 120:741 - 51; http://dx.doi.org/10.1111/j.1471-4159.2011.07592.x; PMID: 22118475
- Tanaka Y, Irie K, Hirota T, Sakisaka T, Nakanishi H, Takai Y. Ectodomain shedding of nectin-1α by SF/HGF and TPA in MDCK cells. Biochem Biophys Res Commun 2002; 299:472 - 8; http://dx.doi.org/10.1016/S0006-291X(02)02681-5; PMID: 12445825
- Fabre-Lafay S, Garrido-Urbani S, Reymond N, Gonçalves A, Dubreuil P, Lopez M. Nectin-4, a new serological breast cancer marker, is a substrate for tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17. J Biol Chem 2005; 280:19543 - 50; http://dx.doi.org/10.1074/jbc.M410943200; PMID: 15784625
- Simpson CL, Kojima S, Cooper-Whitehair V, Getsios S, Green KJ. Plakoglobin rescues adhesive defects induced by ectodomain truncation of the desmosomal cadherin desmoglein 1: implications for exfoliative toxin-mediated skin blistering. Am J Pathol 2010; 177:2921 - 37; http://dx.doi.org/10.2353/ajpath.2010.100397; PMID: 21075858
- Jiang R, Shi Z, Johnson JJ, Liu Y, Stack MS. Kallikrein-5 promotes cleavage of desmoglein-1 and loss of cell-cell cohesion in oral squamous cell carcinoma. J Biol Chem 2011; 286:9127 - 35; http://dx.doi.org/10.1074/jbc.M110.191361; PMID: 21163944
- Getsios S, Simpson CL, Kojima S, Harmon R, Sheu LJ, Dusek RL, et al. Desmoglein 1-dependent suppression of EGFR signaling promotes epidermal differentiation and morphogenesis. J Cell Biol 2009; 185:1243 - 58; http://dx.doi.org/10.1083/jcb.200809044; PMID: 19546243
- Amagai M, Matsuyoshi N, Wang ZH, Andl C, Stanley JR. Toxin in bullous impetigo and staphylococcal scalded-skin syndrome targets desmoglein 1. Nat Med 2000; 6:1275 - 7; http://dx.doi.org/10.1038/81385; PMID: 11062541
- Hanakawa Y, Schechter NM, Lin C, Garza L, Li H, Yamaguchi T, et al. Molecular mechanisms of blister formation in bullous impetigo and staphylococcal scalded skin syndrome. J Clin Invest 2002; 110:53 - 60; PMID: 12093888
- Yang T, Liang D, Koch PJ, Hohl D, Kheradmand F, Overbeek PA. Epidermal detachment, desmosomal dissociation, and destabilization of corneodesmosin in Spink5-/- mice. Genes Dev 2004; 18:2354 - 8; http://dx.doi.org/10.1101/gad.1232104; PMID: 15466487
- Descargues P, Deraison C, Bonnart C, Kreft M, Kishibe M, Ishida-Yamamoto A, et al. Spink5-deficient mice mimic Netherton syndrome through degradation of desmoglein 1 by epidermal protease hyperactivity. Nat Genet 2005; 37:56 - 65; PMID: 15619623
- Wadhawan V, Kolhe YA, Sangith N, Gautam AK, Venkatraman P. From prediction to experimental validation: desmoglein 2 is a functionally relevant substrate of matriptase in epithelial cells and their reciprocal relationship is important for cell adhesion. Biochem J 2012; 447:61 - 70; http://dx.doi.org/10.1042/BJ20111432; PMID: 22783993
- Bech-Serra JJ, Santiago-Josefat B, Esselens C, Saftig P, Baselga J, Arribas J, et al. Proteomic identification of desmoglein 2 and activated leukocyte cell adhesion molecule as substrates of ADAM17 and ADAM10 by difference gel electrophoresis. Mol Cell Biol 2006; 26:5086 - 95; http://dx.doi.org/10.1128/MCB.02380-05; PMID: 16782893
- Sternlicht MD, Werb Z. How matrix metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol 2001; 17:463 - 516; http://dx.doi.org/10.1146/annurev.cellbio.17.1.463; PMID: 11687497
- Bourboulia D, Stetler-Stevenson WG. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): Positive and negative regulators in tumor cell adhesion. Semin Cancer Biol 2010; 20:161 - 8; http://dx.doi.org/10.1016/j.semcancer.2010.05.002; PMID: 20470890
- Wheelock MJ, Buck CA, Bechtol KB, Damsky CH. Soluble 80-kd fragment of cell-CAM 120/80 disrupts cell-cell adhesion. J Cell Biochem 1987; 34:187 - 202; http://dx.doi.org/10.1002/jcb.240340305; PMID: 3611200
- Ryniers F, Stove C, Goethals M, Brackenier L, Noë V, Bracke M, et al. Plasmin produces an E-cadherin fragment that stimulates cancer cell invasion. Biol Chem 2002; 383:159 - 65; http://dx.doi.org/10.1515/BC.2002.016; PMID: 11928810
- Johnson SK, Ramani VC, Hennings L, Haun RS. Kallikrein 7 enhances pancreatic cancer cell invasion by shedding E-cadherin. Cancer 2007; 109:1811 - 20; http://dx.doi.org/10.1002/cncr.22606; PMID: 17354228
- Afonso AA, Sobrin L, Monroy DC, Selzer M, Lokeshwar B, Pflugfelder SC. Tear fluid gelatinase B activity correlates with IL-1α concentration and fluorescein clearance in ocular rosacea. Invest Ophthalmol Vis Sci 1999; 40:2506 - 12; PMID: 10509643
- Smith VA, Rishmawi H, Hussein H, Easty DL. Tear film MMP accumulation and corneal disease. Br J Ophthalmol 2001; 85:147 - 53; http://dx.doi.org/10.1136/bjo.85.2.147; PMID: 11159476
- Pflugfelder SC, Farley W, Luo L, Chen LZ, de Paiva CS, Olmos LC, et al. Matrix metalloproteinase-9 knockout confers resistance to corneal epithelial barrier disruption in experimental dry eye. Am J Pathol 2005; 166:61 - 71; http://dx.doi.org/10.1016/S0002-9440(10)62232-8; PMID: 15632000
- Wong V, Gumbiner BM. A synthetic peptide corresponding to the extracellular domain of occludin perturbs the tight junction permeability barrier. J Cell Biol 1997; 136:399 - 409; http://dx.doi.org/10.1083/jcb.136.2.399; PMID: 9015310
- Makagiansar IT, Avery M, Hu Y, Audus KL, Siahaan TJ. Improving the selectivity of HAV-peptides in modulating E-cadherin-E-cadherin interactions in the intercellular junction of MDCK cell monolayers. Pharm Res 2001; 18:446 - 53; http://dx.doi.org/10.1023/A:1011094025008; PMID: 11451030
- Nava P, López S, Arias CF, Islas S, González-Mariscal L. The rotavirus surface protein VP8 modulates the gate and fence function of tight junctions in epithelial cells. J Cell Sci 2004; 117:5509 - 19; http://dx.doi.org/10.1242/jcs.01425; PMID: 15494377
- Mrsny RJ, Brown GT, Gerner-Smidt K, Buret AG, Meddings JB, Quan C, et al. A key claudin extracellular loop domain is critical for epithelial barrier integrity. Am J Pathol 2008; 172:905 - 15; http://dx.doi.org/10.2353/ajpath.2008.070698; PMID: 18349130
- Beyer I, van Rensburg R, Strauss R, Li Z, Wang H, Persson J, et al. Epithelial junction opener JO-1 improves monoclonal antibody therapy of cancer. Cancer Res 2011; 71:7080 - 90; http://dx.doi.org/10.1158/0008-5472.CAN-11-2009; PMID: 21990319
- Bendas G, Borsig L. Cancer cell adhesion and metastasis: selectins, integrins, and the inhibitory potential of heparins. Int J Cell Biol 2012; 2012:676731; http://dx.doi.org/10.1155/2012/676731; PMID: 22505933
- Lochter A, Galosy S, Muschler J, Freedman N, Werb Z, Bissell MJ. Matrix metalloproteinase stromelysin-1 triggers a cascade of molecular alterations that leads to stable epithelial-to-mesenchymal conversion and a premalignant phenotype in mammary epithelial cells. J Cell Biol 1997; 139:1861 - 72; http://dx.doi.org/10.1083/jcb.139.7.1861; PMID: 9412478
- Xian W, Schwertfeger KL, Vargo-Gogola T, Rosen JM. Pleiotropic effects of FGFR1 on cell proliferation, survival, and migration in a 3D mammary epithelial cell model. J Cell Biol 2005; 171:663 - 73; http://dx.doi.org/10.1083/jcb.200505098; PMID: 16301332
- Brouxhon SM, Kyrkanides S, Teng X, Athar M, Ghazizadeh S, Simon M, et al. Soluble E-cadherin: a critical oncogene modulating receptor tyrosine kinases, MAPK and PI3K/Akt/mTOR signaling. Oncogene 2013; http://dx.doi.org/10.1038/onc.2012.563; PMID: 23318419
- Nawrocki-Raby B, Gilles C, Polette M, Bruyneel E, Laronze JY, Bonnet N, et al. Upregulation of MMPs by soluble E-cadherin in human lung tumor cells. Int J Cancer 2003; 105:790 - 5; http://dx.doi.org/10.1002/ijc.11168; PMID: 12767064
- Najy AJ, Day KC, Day ML. The ectodomain shedding of E-cadherin by ADAM15 supports ErbB receptor activation. J Biol Chem 2008; 283:18393 - 401; http://dx.doi.org/10.1074/jbc.M801329200; PMID: 18434311
- Fouquet S, Lugo-Martínez VH, Faussat AM, Renaud F, Cardot P, Chambaz J, et al. Early loss of E-cadherin from cell-cell contacts is involved in the onset of Anoikis in enterocytes. J Biol Chem 2004; 279:43061 - 9; http://dx.doi.org/10.1074/jbc.M405095200; PMID: 15292248
- Lanza A, Cirillo N. Caspase-dependent cleavage of desmoglein 1 depends on the apoptotic stimulus. Br J Dermatol 2007; 156:400 - 2; http://dx.doi.org/10.1111/j.1365-2133.2006.07654.x; PMID: 17223900
- Steinhusen U, Weiske J, Badock V, Tauber R, Bommert K, Huber O. Cleavage and shedding of E-cadherin after induction of apoptosis. J Biol Chem 2001; 276:4972 - 80; http://dx.doi.org/10.1074/jbc.M006102200; PMID: 11076937
- Rios-Doria J, Day ML. Truncated E-cadherin potentiates cell death in prostate epithelial cells. Prostate 2005; 63:259 - 68; http://dx.doi.org/10.1002/pros.20179; PMID: 15538719
- Sumitomo T, Nakata M, Higashino M, Jin Y, Terao Y, Fujinaga Y, et al. Streptolysin S contributes to group A streptococcal translocation across an epithelial barrier. J Biol Chem 2011; 286:2750 - 61; http://dx.doi.org/10.1074/jbc.M110.171504; PMID: 21084306
- Chun J, Prince A. TLR2-induced calpain cleavage of epithelial junctional proteins facilitates leukocyte transmigration. Cell Host Microbe 2009; 5:47 - 58; http://dx.doi.org/10.1016/j.chom.2008.11.009; PMID: 19154987
- Ito K, Okamoto I, Araki N, Kawano Y, Nakao M, Fujiyama S, et al. Calcium influx triggers the sequential proteolysis of extracellular and cytoplasmic domains of E-cadherin, leading to loss of β-catenin from cell-cell contacts. Oncogene 1999; 18:7080 - 90; http://dx.doi.org/10.1038/sj.onc.1203191; PMID: 10597309
- Cirillo N, Lanza M, De Rosa A, Cammarota M, La Gatta A, Gombos F, et al. The most widespread desmosomal cadherin, desmoglein 2, is a novel target of caspase 3-mediated apoptotic machinery. J Cell Biochem 2008; 103:598 - 606; http://dx.doi.org/10.1002/jcb.21431; PMID: 17559062
- Kolegraff K, Nava P, Laur O, Parkos CA, Nusrat A. Characterization of full-length and proteolytic cleavage fragments of desmoglein-2 in native human colon and colonic epithelial cell lines. Cell Adh Migr 2011; 5:306 - 14; http://dx.doi.org/10.4161/cam.5.4.16911; PMID: 21715983
- Zuckerman JD, Lee WY, DelGaudio JM, Moore CE, Nava P, Nusrat A, et al. Pathophysiology of nasal polyposis: the role of desmosomal junctions. Am J Rhinol 2008; 22:589 - 97; http://dx.doi.org/10.2500/ajr.2008.22.3235; PMID: 19178795
- Brennan D, Peltonen S, Dowling A, Medhat W, Green KJ, Wahl JK 3rd, et al. A role for caveolin-1 in desmoglein binding and desmosome dynamics. Oncogene 2012; 31:1636 - 48; http://dx.doi.org/10.1038/onc.2011.346; PMID: 21841821
- Brennan D, Hu Y, Joubeh S, Choi YW, Whitaker-Menezes D, O’Brien T, et al. Suprabasal Dsg2 expression in transgenic mouse skin confers a hyperproliferative and apoptosis-resistant phenotype to keratinocytes. J Cell Sci 2007; 120:758 - 71; http://dx.doi.org/10.1242/jcs.03392; PMID: 17284515
- Raurell I, Codina M, Casagolda D, Del Valle B, Baulida J, de Herreros AG, et al. Gamma-secretase-dependent and -independent effects of presenilin1 on β-catenin.Tcf-4 transcriptional activity. PLoS One 2008; 3:e4080; http://dx.doi.org/10.1371/journal.pone.0004080; PMID: 19114997
- Ferber EC, Kajita M, Wadlow A, Tobiansky L, Niessen C, Ariga H, et al. A role for the cleaved cytoplasmic domain of E-cadherin in the nucleus. J Biol Chem 2008; 283:12691 - 700; http://dx.doi.org/10.1074/jbc.M708887200; PMID: 18356166
- Jang C, Choi JK, Na YJ, Jang B, Wasco W, Buxbaum JD, et al. Calsenilin regulates presenilin 1/γ-secretase-mediated N-cadherin ε-cleavage and β-catenin signaling. FASEB J 2011; 25:4174 - 83; http://dx.doi.org/10.1096/fj.11-185926; PMID: 21852538
- Maetzel D, Denzel S, Mack B, Canis M, Went P, Benk M, et al. Nuclear signalling by tumour-associated antigen EpCAM. Nat Cell Biol 2009; 11:162 - 71; http://dx.doi.org/10.1038/ncb1824; PMID: 19136966
- Utton MA, Eickholt B, Howell FV, Wallis J, Doherty P. Soluble N-cadherin stimulates fibroblast growth factor receptor dependent neurite outgrowth and N-cadherin and the fibroblast growth factor receptor co-cluster in cells. J Neurochem 2001; 76:1421 - 30; http://dx.doi.org/10.1046/j.1471-4159.2001.00140.x; PMID: 11238727
- Reddy P, Liu L, Ren C, Lindgren P, Boman K, Shen Y, et al. Formation of E-cadherin-mediated cell-cell adhesion activates AKT and mitogen activated protein kinase via phosphatidylinositol 3 kinase and ligand-independent activation of epidermal growth factor receptor in ovarian cancer cells. Mol Endocrinol 2005; 19:2564 - 78; http://dx.doi.org/10.1210/me.2004-0342; PMID: 15928314
- Lampugnani MG, Orsenigo F, Gagliani MC, Tacchetti C, Dejana E. Vascular endothelial cadherin controls VEGFR-2 internalization and signaling from intracellular compartments. J Cell Biol 2006; 174:593 - 604; http://dx.doi.org/10.1083/jcb.200602080; PMID: 16893970
- Kang HG, Jenabi JM, Zhang J, Keshelava N, Shimada H, May WA, et al. E-cadherin cell-cell adhesion in ewing tumor cells mediates suppression of anoikis through activation of the ErbB4 tyrosine kinase. Cancer Res 2007; 67:3094 - 105; http://dx.doi.org/10.1158/0008-5472.CAN-06-3259; PMID: 17409416
- Pittard AJ, Banks RE, Galley HF, Webster NR. Soluble E-cadherin concentrations in patients with systemic inflammatory response syndrome and multiorgan dysfunction syndrome. Br J Anaesth 1996; 76:629 - 31; http://dx.doi.org/10.1093/bja/76.5.629; PMID: 8688260
- Katayama M, Hirai S, Kamihagi K, Nakagawa K, Yasumoto M, Kato I. Soluble E-cadherin fragments increased in circulation of cancer patients. Br J Cancer 1994; 69:580 - 5; http://dx.doi.org/10.1038/bjc.1994.106; PMID: 8123491
- Sewpaul A, French JJ, Khoo TK, Kernohan M, Kirby JA, Charnley RM. Soluble E-cadherin: an early marker of severity in acute pancreatitis. HPB Surg 2009; 2009:397375; http://dx.doi.org/10.1155/2009/397375; PMID: 19421334
- Doyle JL, Haas TL. Differential role of beta-catenin in VEGF and histamine-induced MMP-2 production in microvascular endothelial cells. J Cell Biochem 2009; 107:272 - 83; http://dx.doi.org/10.1002/jcb.22123; PMID: 19306293
- He W, Tan RJ, Li Y, Wang D, Nie J, Hou FF, et al. Matrix metalloproteinase-7 as a surrogate marker predicts renal Wnt/β-catenin activity in CKD. J Am Soc Nephrol 2012; 23:294 - 304; http://dx.doi.org/10.1681/ASN.2011050490; PMID: 22095947
- van Kempen LC, de Visser KE, Coussens LM. Inflammation, proteases and cancer. Eur J Cancer 2006; 42:728 - 34; http://dx.doi.org/10.1016/j.ejca.2006.01.004; PMID: 16524717
- Chung Y, Law S, Kwong DL, Luk JM. Serum soluble E-cadherin is a potential prognostic marker in esophageal squamous cell carcinoma. Dis Esophagus 2011; 24:49 - 55; http://dx.doi.org/10.1111/j.1442-2050.2010.01093.x; PMID: 20807231
- Derycke L, De Wever O, Stove V, Vanhoecke B, Delanghe J, Depypere H, et al. Soluble N-cadherin in human biological fluids. Int J Cancer 2006; 119:2895 - 900; http://dx.doi.org/10.1002/ijc.22219; PMID: 16998833
- Griffiths TR, Brotherick I, Bishop RI, White MD, McKenna DM, Horne CH, et al. Cell adhesion molecules in bladder cancer: soluble serum E-cadherin correlates with predictors of recurrence. Br J Cancer 1996; 74:579 - 84; http://dx.doi.org/10.1038/bjc.1996.404; PMID: 8761373
- Protheroe AS, Banks RE, Mzimba M, Porter WH, Southgate J, Singh PN, et al. Urinary concentrations of the soluble adhesion molecule E-cadherin and total protein in patients with bladder cancer. Br J Cancer 1999; 80:273 - 8; http://dx.doi.org/10.1038/sj.bjc.6690351; PMID: 10390008
- Wilmanns C, Grossmann J, Steinhauer S, Manthey G, Weinhold B, Schmitt-Gräff A, et al. Soluble serum E-cadherin as a marker of tumour progression in colorectal cancer patients. Clin Exp Metastasis 2004; 21:75 - 8; http://dx.doi.org/10.1023/B:CLIN.0000017204.38807.22; PMID: 15065605
- Weiss JV, Klein-Scory S, Kübler S, Reinacher-Schick A, Stricker I, Schmiegel W, et al. Soluble E-cadherin as a serum biomarker candidate: elevated levels in patients with late-stage colorectal carcinoma and FAP. Int J Cancer 2011; 128:1384 - 92; http://dx.doi.org/10.1002/ijc.25438; PMID: 20473926
- Chan AO, Lam SK, Chu KM, Lam CM, Kwok E, Leung SY, et al. Soluble E-cadherin is a valid prognostic marker in gastric carcinoma. Gut 2001; 48:808 - 11; http://dx.doi.org/10.1136/gut.48.6.808; PMID: 11358900
- Soyama A, Eguchi S, Takatsuki M, Kawashita Y, Hidaka M, Tokai H, et al. Significance of the serum level of soluble E-cadherin in patients with HCC. Hepatogastroenterology 2008; 55:1390 - 3; PMID: 18795696
- Charalabopoulos K, Gogali A, Dalavaga Y, Daskalopoulos G, Vassiliou M, Bablekos G, et al. The clinical significance of soluble E-cadherin in nonsmall cell lung cancer. Exp Oncol 2006; 28:83 - 5; PMID: 16614715
- Gogali A, Charalabopoulos K, Zampira I, Konstantinidis AK, Tachmazoglou F, Daskalopoulos G, et al. Soluble adhesion molecules E-cadherin, intercellular adhesion molecule-1, and E-selectin as lung cancer biomarkers. Chest 2010; 138:1173 - 9; http://dx.doi.org/10.1378/chest.10-0157; PMID: 20495107
- Sundfeldt K, Ivarsson K, Rask K, Haeger M, Hedin L, Brännström M. Higher levels of soluble E-cadherin in cyst fluid from malignant ovarian tumours than in benign cysts. Anticancer Res 2001; 21:1A 65 - 70; PMID: 11299791
- Gil OD, Lee C, Ariztia EV, Wang FQ, Smith PJ, Hope JM, et al. Lysophosphatidic acid (LPA) promotes E-cadherin ectodomain shedding and OVCA429 cell invasion in an uPA-dependent manner. Gynecol Oncol 2008; 108:361 - 9; http://dx.doi.org/10.1016/j.ygyno.2007.10.027; PMID: 18073130
- Kuefer R, Hofer MD, Zorn CS, Engel O, Volkmer BG, Juarez-Brito MA, et al. Assessment of a fragment of e-cadherin as a serum biomarker with predictive value for prostate cancer. Br J Cancer 2005; 92:2018 - 23; http://dx.doi.org/10.1038/sj.bjc.6602599; PMID: 15870707
- Billion K, Ibrahim H, Mauch C, Niessen CM. Increased soluble E-cadherin in melanoma patients. Skin Pharmacol Physiol 2006; 19:65 - 70; http://dx.doi.org/10.1159/000091972; PMID: 16685144
- Shirahama S, Furukawa F, Wakita H, Takigawa M. E- and P-cadherin expression in tumor tissues and soluble E-cadherin levels in sera of patients with skin cancer. J Dermatol Sci 1996; 13:30 - 6; http://dx.doi.org/10.1016/0923-1811(95)00493-9; PMID: 8902651
- Velikova G, Banks RE, Gearing A, Hemingway I, Forbes MA, Preston SR, et al. Serum concentrations of soluble adhesion molecules in patients with colorectal cancer. Br J Cancer 1998; 77:1857 - 63; http://dx.doi.org/10.1038/bjc.1998.309; PMID: 9667659
- Gadducci A, Ferdeghini M, Cosio S, Annicchiarico C, Ciampi B, Bianchi R, et al. Preoperative serum E-cadherin assay in patients with ovarian carcinoma. Anticancer Res 1999; 19:1B 769 - 72; PMID: 10216491
- Manetti M, Guiducci S, Romano E, Rosa I, Ceccarelli C, Mello T, et al. Differential expression of junctional adhesion molecules in different stages of systemic sclerosis. Arthritis Rheum 2013; 65:247 - 57; http://dx.doi.org/10.1002/art.37712; PMID: 23001478
- Streeck H, Kwon DS, Pyo A, Flanders M, Chevalier MF, Law K, et al. Epithelial adhesion molecules can inhibit HIV-1-specific CD8⁺ T-cell functions. Blood 2011; 117:5112 - 22; http://dx.doi.org/10.1182/blood-2010-12-321588; PMID: 21403126
- Nishifuji K, Shimizu A, Ishiko A, Iwasaki T, Amagai M. Removal of amino-terminal extracellular domains of desmoglein 1 by staphylococcal exfoliative toxin is sufficient to initiate epidermal blister formation. J Dermatol Sci 2010; 59:184 - 91; http://dx.doi.org/10.1016/j.jdermsci.2010.07.010; PMID: 20728315
- Schnell U, Kuipers J, Giepmans BN. EpCAM proteolysis: new fragments with distinct functions?. Biosci Rep 2013; 33:e00030; http://dx.doi.org/10.1042/BSR20120128; PMID: 23409978
- Casas E, Barron C, Francis SA, McCormack JM, McCarthy KM, Schneeberger EE, et al. Cholesterol efflux stimulates metalloproteinase-mediated cleavage of occludin and release of extracellular membrane particles containing its C-terminal fragments. Exp Cell Res 2010; 316:353 - 65; http://dx.doi.org/10.1016/j.yexcr.2009.10.020; PMID: 19854171
- Zeng R, Li X, Gorodeski GI. Estrogen abrogates transcervical tight junctional resistance by acceleration of occludin modulation. J Clin Endocrinol Metab 2004; 89:5145 - 55; http://dx.doi.org/10.1210/jc.2004-0823; PMID: 15472219
- Gorodeski GI. Estrogen decrease in tight junctional resistance involves matrix-metalloproteinase-7-mediated remodeling of occludin. Endocrinology 2007; 148:218 - 31; http://dx.doi.org/10.1210/en.2006-1120; PMID: 17038551
- Biswas MH, Du C, Zhang C, Straubhaar J, Languino LR, Balaji KC. Protein kinase D1 inhibits cell proliferation through matrix metalloproteinase-2 and matrix metalloproteinase-9 secretion in prostate cancer. Cancer Res 2010; 70:2095 - 104; http://dx.doi.org/10.1158/0008-5472.CAN-09-4155; PMID: 20160036
- Lynch CC, Vargo-Gogola T, Matrisian LM, Fingleton B. Cleavage of E-Cadherin by Matrix Metalloproteinase-7 Promotes Cellular Proliferation in Nontransformed Cell Lines via Activation of RhoA. J Oncol 2010; 2010:530745; http://dx.doi.org/10.1155/2010/530745; PMID: 20628524
- McGuire JK, Li Q, Parks WC. Matrilysin (matrix metalloproteinase-7) mediates E-cadherin ectodomain shedding in injured lung epithelium. Am J Pathol 2003; 162:1831 - 43; http://dx.doi.org/10.1016/S0002-9440(10)64318-0; PMID: 12759241
- Lee KH, Choi EY, Hyun MS, Jang BI, Kim TN, Kim SW, et al. Association of extracellular cleavage of E-cadherin mediated by MMP-7 with HGF-induced in vitro invasion in human stomach cancer cells. Eur Surg Res 2007; 39:208 - 15; http://dx.doi.org/10.1159/000101452; PMID: 17396032
- Zuo JH, Zhu W, Li MY, Li XH, Yi H, Zeng GQ, et al. Activation of EGFR promotes squamous carcinoma SCC10A cell migration and invasion via inducing EMT-like phenotype change and MMP-9-mediated degradation of E-cadherin. J Cell Biochem 2011; 112:2508 - 17; http://dx.doi.org/10.1002/jcb.23175; PMID: 21557297
- Huguenin M, Müller EJ, Trachsel-Rösmann S, Oneda B, Ambort D, Sterchi EE, et al. The metalloprotease meprinbeta processes E-cadherin and weakens intercellular adhesion. PLoS One 2008; 3:e2153; http://dx.doi.org/10.1371/journal.pone.0002153; PMID: 18478055
- Maretzky T, Scholz F, Köten B, Proksch E, Saftig P, Reiss K. ADAM10-mediated E-cadherin release is regulated by proinflammatory cytokines and modulates keratinocyte cohesion in eczematous dermatitis. J Invest Dermatol 2008; 128:1737 - 46; http://dx.doi.org/10.1038/sj.jid.5701242; PMID: 18200054
- Schirrmeister W, Gnad T, Wex T, Higashiyama S, Wolke C, Naumann M, et al. Ectodomain shedding of E-cadherin and c-Met is induced by Helicobacter pylori infection. Exp Cell Res 2009; 315:3500 - 8; http://dx.doi.org/10.1016/j.yexcr.2009.07.029; PMID: 19665015
- Solanas G, Cortina C, Sevillano M, Batlle E. Cleavage of E-cadherin by ADAM10 mediates epithelial cell sorting downstream of EphB signalling. Nat Cell Biol 2011; 13:1100 - 7; http://dx.doi.org/10.1038/ncb2298; PMID: 21804545
- Frank CF, Hostetter MK. Cleavage of E-cadherin: a mechanism for disruption of the intestinal epithelial barrier by Candida albicans. Transl Res 2007; 149:211 - 22; http://dx.doi.org/10.1016/j.trsl.2006.11.006; PMID: 17383595
- Klucky B, Mueller R, Vogt I, Teurich S, Hartenstein B, Breuhahn K, et al. Kallikrein 6 induces E-cadherin shedding and promotes cell proliferation, migration, and invasion. Cancer Res 2007; 67:8198 - 206; http://dx.doi.org/10.1158/0008-5472.CAN-07-0607; PMID: 17804733
- Mayerle J, Schnekenburger J, Krüger B, Kellermann J, Ruthenbürger M, Weiss FU, et al. Extracellular cleavage of E-cadherin by leukocyte elastase during acute experimental pancreatitis in rats. Gastroenterology 2005; 129:1251 - 67; http://dx.doi.org/10.1053/j.gastro.2005.08.002; PMID: 16230078
- Gocheva V, Zeng W, Ke D, Klimstra D, Reinheckel T, Peters C, et al. Distinct roles for cysteine cathepsin genes in multistage tumorigenesis. Genes Dev 2006; 20:543 - 56; http://dx.doi.org/10.1101/gad.1407406; PMID: 16481467
- Dusek RL, Getsios S, Chen F, Park JK, Amargo EV, Cryns VL, et al. The differentiation-dependent desmosomal cadherin desmoglein 1 is a novel caspase-3 target that regulates apoptosis in keratinocytes. J Biol Chem 2006; 281:3614 - 24; http://dx.doi.org/10.1074/jbc.M508258200; PMID: 16286477
- Lundström A, Egelrud T. Evidence that cell shedding from plantar stratum corneum in vitro involves endogenous proteolysis of the desmosomal protein desmoglein I. J Invest Dermatol 1990; 94:216 - 20; http://dx.doi.org/10.1111/1523-1747.ep12874531; PMID: 1688905
- Klessner JL, Desai BV, Amargo EV, Getsios S, Green KJ. EGFR and ADAMs cooperate to regulate shedding and endocytic trafficking of the desmosomal cadherin desmoglein 2. Mol Biol Cell 2009; 20:328 - 37; http://dx.doi.org/10.1091/mbc.E08-04-0356; PMID: 18987342
- Santiago-Josefat B, Esselens C, Bech-Serra JJ, Arribas J. Post-transcriptional up-regulation of ADAM17 upon epidermal growth factor receptor activation and in breast tumors. J Biol Chem 2007; 282:8325 - 31; http://dx.doi.org/10.1074/jbc.M608826200; PMID: 17227756
- Ramani VC, Hennings L, Haun RS. Desmoglein 2 is a substrate of kallikrein 7 in pancreatic cancer. BMC Cancer 2008; 8:373; http://dx.doi.org/10.1186/1471-2407-8-373; PMID: 19091121
- Weiske J, Schöneberg T, Schröder W, Hatzfeld M, Tauber R, Huber O. The fate of desmosomal proteins in apoptotic cells. J Biol Chem 2001; 276:41175 - 81; http://dx.doi.org/10.1074/jbc.M105769200; PMID: 11500511
- Covington MD, Burghardt RC, Parrish AR. Ischemia-induced cleavage of cadherins in NRK cells requires MT1-MMP (MMP-14). Am J Physiol Renal Physiol 2006; 290:F43 - 51; http://dx.doi.org/10.1152/ajprenal.00179.2005; PMID: 16077081
- Yoo CB, Yun SM, Jo C, Koh YH. γ-Secretase-dependent cleavage of E-cadherin by staurosporine in breast cancer cells. Cell Commun Adhes 2012; 19:11 - 6; http://dx.doi.org/10.3109/15419061.2012.665969; PMID: 22401168
- Schmeiser K, Grand RJ. The fate of E- and P-cadherin during the early stages of apoptosis. Cell Death Differ 1999; 6:377 - 86; http://dx.doi.org/10.1038/sj.cdd.4400504; PMID: 10381631
- Kim DY, Ingano LA, Kovacs DM. Nectin-1α, an immunoglobulin-like receptor involved in the formation of synapses, is a substrate for presenilin/γ-secretase-like cleavage. J Biol Chem 2002; 277:49976 - 81; http://dx.doi.org/10.1074/jbc.M210179200; PMID: 12376527