Abstract
There are three stages of transcription: initiation, elongation and termination, and traditionally there has been a clear distinction between the stages. The specificity factor sigma is completely released from bacterial RNA polymerase after initiation, and then recycled for another round of transcription. Elongation factors then associate with the polymerase followed by termination factors (where necessary). These factors dissociate prior to initiation of a new round of transcription. However, there is growing evidence suggesting that sigma factors can be retained in the elongation complex. The structure of bacterial RNAP in complex with an essential elongation factor NusA has recently been published, which suggested rather than competing for the major σ binding site, NusA binds to a discrete region on RNAP. A model was proposed to help explain the way in which both factors could be associated with RNAP during the transition from transcription initiation to elongation.
Acknowledgements
Work in the lab of P.L. is supported by the ARC and NHMRC. X.Y. was supported by an Australian Postgraduate Award from the Australian Government.
Figures and Tables
Figure 1 (A) Structural model of RNAP with the relative positions of the α, β, β' and ω subunits marked. The boxed areas indicate the clamp-helix (CH) region and β-flaps which are shown expanded below. σ region 2 binds the CH region, whilst σ region 4 and NusA NTD bind the β-flap. (B–E) Cartoon representation of the transcription machinery at different stages. (B) RNAP scans along the DNA until it encounters a promoter. (C) After a few rounds of abortive initiation, σ factor region 4 dissociates from the β flap, while region 2 remains bound to the CH region. (D) NusA binds to RNAP via its NTD while the rest of σ remains associated. (E) In most cases, σ is released from RNAP, resulting in a stable EC. See main text for detailed discussion.
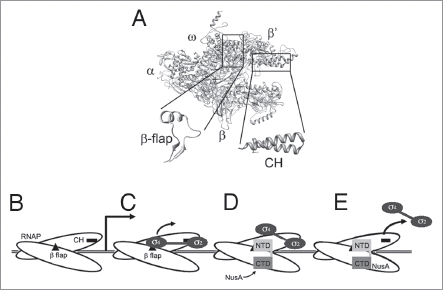
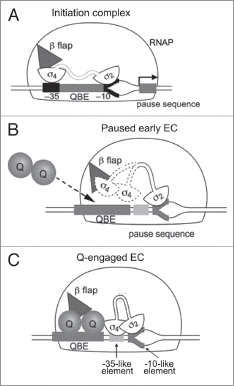
Figure 2 Formation of the phage Q protein antitermination complex at the PR' promoter. (A) The initiation complex with 2 bound to the −10 promoter element and 4 to the −35 promoter element. 4 is also shown interacting with the β flap (triangle) of RNAP. (B) The paused early EC at PR' with 2 bound to the pause-inducing −10-like element. 4 (dotted outline) is released from the β flap. (C) Q (shown as a dimer) binds to the QBE and become associated with the paused EC at the β flap region. Q also interacts with 4 and stabilizes its binding to the −35-like pausing element. Adapted from Nickels et al (ref. Citation16). See main text for detailed discussion.