
Abstract
It has been recently stated that stress-responding genes in yeast are enriched in cryptic transcripts and that this is the cause of the differences observed between mRNA amount and RNA polymerase occupancy profiles. Other studies have shown that such differences are mainly due to modulation of mRNA stabilities. Here we analyze the relationship between the presence of cryptic transcripts in genes and their stress response profiles. Despite some of the stress-responding gene groups being indeed enriched in specific classes of cryptic transcripts, we found no statistically significant evidence that cryptic transcription is responsible for the differences observed between mRNA and transcription rate profiles.
Acknowledgments
This work has been supported by grants from the Spanish MICINN: BFU2010-17656/BMC to E.H., BFU2010-21975-CO3-01/BMC to J.E.P.O. and BFU2010-21975-CO3-02/BMC to S.C., from the Generalitat Valenciana Prometeo 2011/088 to J.E.P.O. and from Junta de Andalucía (grants P07-CVI02623 and P08-CVI-03508) to S.C. MICINN and Junta de Andalucía grants involve FEDER resources from the European Union. V.P. is supported by an EMBO fellowship.
Note Added in Proof
In a recent paper, Xu et al.Citation23 made an exhaustive study analysis of antisensetranscription in yeast and concluded that, although it affects the regulation of the gene expression, the main driving force of canonical (sense) expression is not its associated antisense expression.
Figures and Tables
Figure 1 Cryptic transcription does not correlate with the differences between the transcription rate and mRNA profiles in heat stress response. We display the kernel density estimators for the r Pearson correlations between the theoretical and experimental mRNA levels in the heat stress response. The estimated densities correspond to genes with cryptic transcripts (dotted line) and to genes without cryptic transcripts (solid line), as determined by Xu et al.Citation17 (A), Neil et al.Citation18 (B) and Yassour et al.Citation2 (C) Note that the same bandwidth has been used for both densities. We have tested the normality hypothesis using the Shapiro-Wilk test and the normal null hypothesis is rejected for all data sets. However, because we have large groups of genes to be compared the means can be compared using al two-sample t test (t, the Welch correction was used). Additionally, the null hypothesis of a common distribution for both groups will be tested using the two-sample Kolmogorov-Smirnov test (K-S). All the statistical tests can be found in reference Citation19. The p values for the K-S test and t tests are indicated in each panel. Statistically significant values are underlined. Theoretical mRNA (RAt) levels were calculated using the equation described in reference 12 and assuming a constant degradation constant (kD) equal to the steady-state value observed before stress (time 0). From that equation, the theoretical mRNA values at different times were calculated from an initial mRNA amount and the experimental TR values.Citation12 Experimental mRNA (RAe) levels were those determined in reference Citation12. r Pearson correlations were calculated for individual gene values of the predicted theoretical mRNA levels plotted vs. the experimental mRNA data. See reference Citation10 and Citation12 for further details of the data sets used.
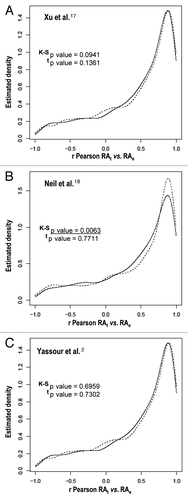
Figure 2 Different correlation patterns in comparisons between cryptic transcription data sets and change ratios after heat stress. We show a representation of the average ratio change (in log2 scale) of each time point after stress in relation to the zero time (x axis) against the density (y axis) of the distribution function for genes with cryptic transcripts (dotted line) and for genes without cryptic transcripts (solid line), as determined by Xu et al.Citation17 (A), Neil et al.Citation18 (B) and Yassour et al.Citation2 (D). The data set of Neil et al.Citation18 is split into sense (C1) and antisense (C2). As in , the distributions and the means of correlation coefficients were tested by using the K-S and t tests, respectively. The p values for the K-S and t tests are indicated in each part. Statistically significant values are underlined.
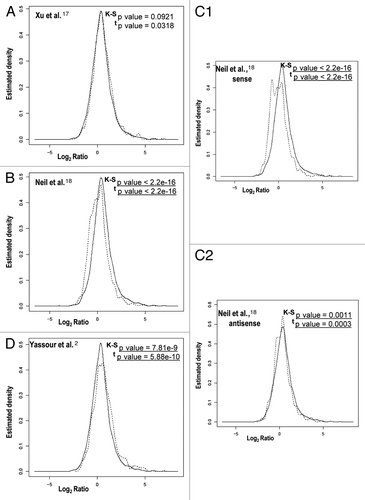
Figure 3 Different cryptic transcription data sets do not significantly overlap. The lists of genes with cryptic transcripts from Xu et al.,Citation17 Neil et al.Citation18 and Yassour et al.Citation2 mostly contain different genes. We used the original data sets but selected only those genes having overlapping cryptic transcripts that were also present in our previous heat stress-response study.Citation12 All the overlappings are not statistically significant when using a hypergeometric test, except for the overlap of Neil et al. and that of Yassour et al.Citation2 which are significantly larger than expected with a p value of 7 × 10−4.
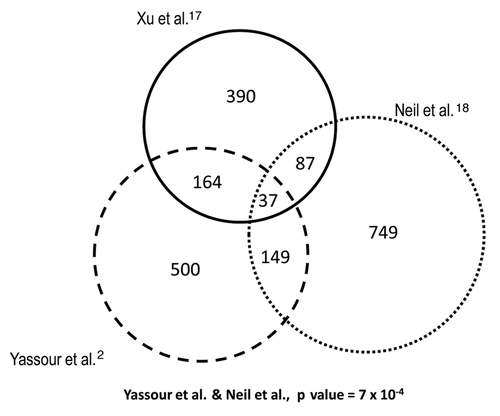
Figure 4 Analyses of up- and downregulated genes after heat stress show a main dependence on the transcription rate change. (A) There is a slight positive dependence of the ratio of change (in log2 with absolute values, either up or downregulated) after stress in relation to the r correlation coefficient, calculated as in . (B) Histogram of the correlation coefficients for the whole set of genes (dotted line) and for two particular groups [96 ribosomal protein (RP) genes (black bars) showing a considerable downregulation and a large proportion of sense cryptic transcription (shown in Fig. S2), and 1,051 upregulated genes (>1.2 on the log2 scale, see Fig. S2; gray bars)]. Note that the scale for the RP genes is magnified ten times.
![Figure 4 Analyses of up- and downregulated genes after heat stress show a main dependence on the transcription rate change. (A) There is a slight positive dependence of the ratio of change (in log2 with absolute values, either up or downregulated) after stress in relation to the r correlation coefficient, calculated as in Figure 1. (B) Histogram of the correlation coefficients for the whole set of genes (dotted line) and for two particular groups [96 ribosomal protein (RP) genes (black bars) showing a considerable downregulation and a large proportion of sense cryptic transcription (shown in Fig. S2), and 1,051 upregulated genes (>1.2 on the log2 scale, see Fig. S2; gray bars)]. Note that the scale for the RP genes is magnified ten times.](/cms/asset/46265cfb-21cc-42fa-9d49-07968c8045d1/ktrn_a_10919416_f0004.gif)