Figures & data
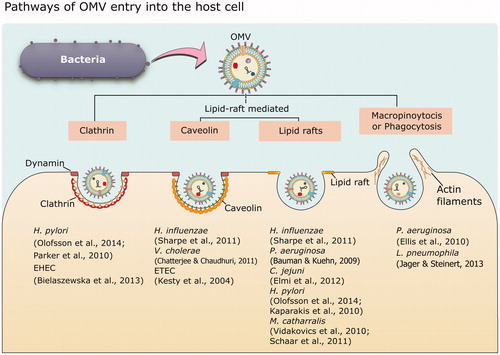
Figure 1. (A) Structure of the gram-negative bacterial cell envelope. The envelope consists of two lipid bilayers and a peptidoglycan layer. The bacterial outer membrane (OM) is unique for its asymmetrical distribution of lipid moieties, wherein the exterior leaflet is enriched in lipopolysaccharides (LPS), and the inner leaflet predominantly contains glycerophospholipid. The inner membrane (IM) is symmetrical in the glycerophospholipid composition in both the leaflets and contains many complex protein assemblies like the YrbDE, Tol component of the Tol-Pal system and NlpA. The region between IM and OM is called as the periplasmic space (PS). The periplasm contains the peptidoglycan (PG) layer along with other protein assemblies such as VacJ/YrbC, Tol B and Pal component of the Tol-Pal system, and Lipoma preferred partner (Lpp). The rest of the Yrb complex components (YrbBF) are juxta-membrane from the cytoplasmic face of IM. (B) The building blocks of the LPS layer. An LPS moiety is composed of the inner and outer core oligosaccharides followed by variable length O-antigens covalently linked to the outer-core. The inner core an LPS moiety contains l-glycerol-d-manno-heptopyranose (l-d-Hep) and Kdo as the major components while the outer core contains d-glucose as the major component linked with l-d-Hep and d-galactose monomers (see text for more details).
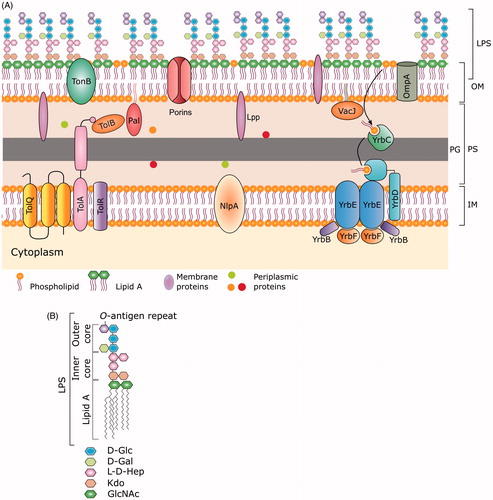
Figure 2. The structure (A) and production of OMVs from gram-negative bacteria (B). The composition of the lipid bilayer of OMVs retains the asymmetry observed in bacterial OM, with complex lipids in outermost layer mounted on a uniform phospholipid inner layer. The outermost layer of OMVs consists of the LPS layer (made up of lipid A, LPS core, and O-antigen) derived from the bacterial OM (see for details of bacterial OM). OMVs host a large repertoire of cargoes such as nucleic acids, enzymes, toxins, fragments of peptidoglycans, and fluid from the periplasmic space. (B) Formation of OMV starts with the bulging of the outer membrane at very small region followed by progress in the expansion of the membrane. The increased expansion is established with the spherical structure of blebs. Eventually vesicle pinches off at the joint neck and gets separated from the cell.
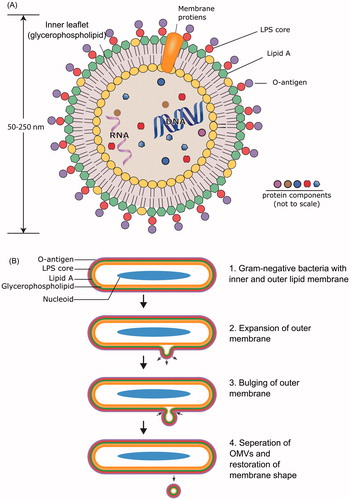
Table 1. Regulatory mechanisms of OMV formation.
Figure 3. Quantitative high-throughput detection of OMVs using scanning ion occlusion spectroscopy (SIOS). Real-time scanning of the conductivity across the pore at different stretches and different pore size allows the discrimination of individual OMV population in a large disperse mixture. (A) Small and (B) large vesicles passing through the pore across the fluid chamber from a to b generate different conductivity signals. Larger vesicles require greater stretching of the pore to increase its diameter. This change in the pore size changes the strength of the current across the fluid chamber and is recorded as a signal of larger amplitude compared to smaller vesicles.
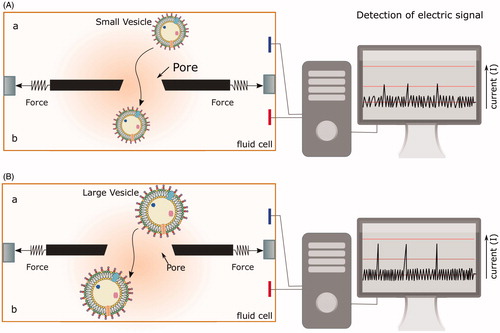
Figure 4. Different modes of OMV entry into the host cell. OMVs can enter the host cells via two major pathways: clathrin-mediated endocytosis, and the lipid raft-mediated pathway. Lipid rafts are dynamic membrane microdomains rich in lipids like cholesterol and sphingolipids, and proteins such as caveolin. Pinocytosis and phagocytosis are other two general pathways involved in uptake of OMVs. The figure provides representative examples of different bacteria that have been shown to enter the host cell exploiting at least one of the above-mentioned pathways.