Figures & data
Figure 1. Feedback paradox I: glucocorticoid feedback resistance evoked by a stressor but not by prior corticosterone. Left: Plasma corticosterone levels resulting from 60 min restraint, sc injections of corticosterone (60 μg) or, sc injection of saline at 0 and 40 min. Right: the responses to injection stress in the three groups are shown. After corticosterone treatment, the response to injection was significantly (p < .01) inhibited compared to either of the other groups. Figure adapted from (Dallman & Jones, Citation1973).
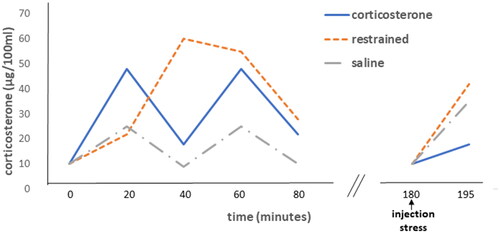
Figure 2. MR- and GR-Mediated actions during information processing. MR and GR mediate in a complementary and balanced manner (see Box 4) the actions of glucocorticoids on the processing of information. MR is promiscuous and its activation by cortisol and corticosterone can be modulated by their 11 deoxy congeners, progesterone, and Aldo. GR is activated at higher concentrations of glucocorticoids after stress and toward/during the activity period in the circadian cycle. The actions exerted by glucocorticoids on information processing are conditional and time-dependent and should be considered in concert with CRH/vasopressin/POMC-driven HPA-axis activity, and sympathetic and cytokine responses together with multiple dedicated signaling cascades. Non-genomic actions rapidly proceed via the lower affinity membrane variants of MR and GR and thus may respond to a wide range of corticosterone concentrations. these rapid actions occur in the domains of arousal. appraisal processes and choice of coping style. The slow genomic responses mediated by MR and GR deal with energy metabolism, adaptation to change, and priming in preparation for the future (memory storage, inflammasome formation, and energy storage). However, the actions of glucocorticoids are also subject to meta-plasticity involving both genomic and non-genomic actions mediated by MR and GR. The actions are modified by receptor coregulator composition and are subject to ultrashort feedback loops via FKBP5. Perception discrimination ability and sensory detection threshold (nociception, taste, smell, hearing) are regulated via GR activation. The appraisal process may frustrate coping if the individual is unable to predict either the outcome of dealing with a threat or due to lack of sufficient energy as a source of motivation to pursue coping with e.g. fear or reward. If the prediction is uncertain the subject arrives in a state of lack of control: the ‘brain gets stuck’ as a consequence of chronic stress and useless energy expenditure (McEwen, 2017; McEwen & Akil, Citation2020; Sousa, Citation2016). The figure depicts an imbalance with excess MR- over GR-mediated actions. Such a positive “MR/GR balance” is characteristic of a sympathetic-driven dominant individual with low circulating corticosterone and a habitual coping style (de Boer et al., Citation2017). In a stable predictable context, this is a healthy resilient phenotype, but during lack of control the vulnerability to stressors is increased (Daskalakis et al., Citation2022; de Kloet et al., Citation1999, Citation2005, Citation2018; de Kloet & Joëls, Citation2023). Blue and red refer to GR- and MR-mediated actions.
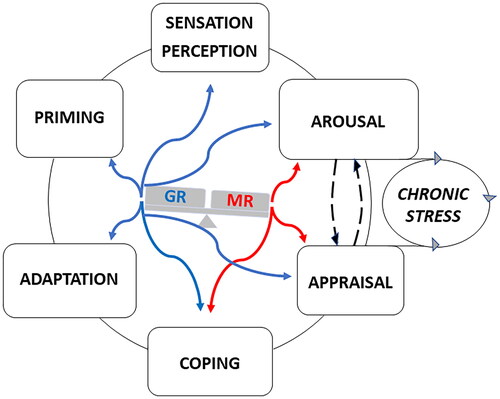
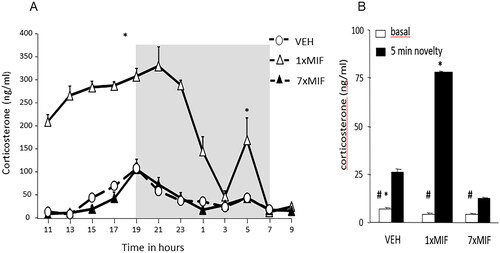
Figure 3. Feedback paradox II: daily GR antagonist mifepristone reveals its apparent agonist action. (a) Circadian secretion of corticosterone in ng/ml measured every 2 h in the blood plasma of male mice C57BL/6J that received mifepristone (MIF) once (1 × 200mg/kg) or for seven days (7 × MIF). Oats + MIF or Oats + VEH were placed in the feeding cup at 0900 h, and consumed within 10 min. Mice were entrained in a 12–12-h light-dark cycle (the dark phase from 1900 to 0700 h represented by the gray-shaded area). (b) Basal and novelty (5 min exposure to the circular hole board)- induced corticosterone (ng/ml) was determined in mice, 24 h after the last administration of VEH, 1 × MIF, or 7 × MIF. Data are presented as mean ± SEM; p < .05* versus other groups, # within groups. Figure adapted from (Dalm et al., Citation2019).