
Figures & data
Figure 1. In vivo and in vitro generation of circRnas.
A. Canonical splicing and back-splicing both refer to distinct processes that a precursor mRNA (pre-mRNA) can undergo. Canonical splicing results in the production of a linear mRNA, while back-splicing leads to the formation of a circRNA and an alternatively spliced linear mRNA variant lacking specific exons. This occurs through a unique process, where the 5’ splice site (5’ ss, donor site) joins with a 3’ splice site (3’ ss, acceptor site) in a ‘head-to-tail’ manner.
B. In the lariat intermediate model, canonical splicing first produces a linear RNA without introns, and a long intron lariat containing the skipped exons. This lariat then undergoes back-splicing, forming a circular RNA.
C. In the Intronic circRNA model, after an intron lariat forms, it can take one of two paths: rapid degradation if stable consensus RNA sequences are absent, or the creation of a circular intronic RNA (ciRNA) when these consensus sequences are present and prevent debranching.
D. T4 RNA ligase facilitates the formation of a covalent 5′,3′-phosphodiester bond by promoting a nucleophilic attack, where the 3′-OH terminus reacts with the activated 5′-terminus. The splint, whether composed of cDNA or RNA, serves as a bridge, assisting in aligning and stabilizing the reactive ends.
E. Group I Intron Self-Splicing System: Attachment of a free guanosine (G-OH) to the half intron initiates transesterification at the 5’ splice site of exon 1. A second transesterification at the 3’ splice site using the freed hydroxyl group at the 3’ end of the intermediate leads to circularization of the inserted sequence and the release of two intron tails.
F. Group II Intron Self-Splicing System: Involves joining the 5′ splice site at the end of an exon to the 3′ splice site at the beginning of the same exon. All exon sequences are substitutable for group II intron catalysed inverse splicing, enabling more accurate linear RNA precursor ligation.
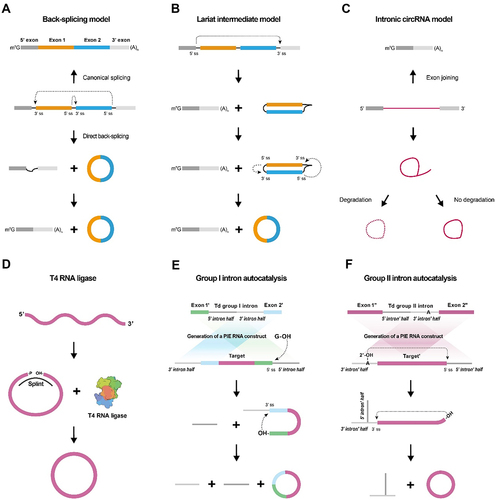
Table 1. The production and function of viral-derived circRNAs.
Figure 2. Perspectives of circRNA nanomedicine.
CircRNA shows promise in various areas including rare disease therapy, CAR-T therapy, vaccine design, antibody expression, cytokine modulation, and diagnostic marker development. It can serve as a target for diagnostic and prognostic tools, and studies indicate its potential in restoring heart function and therapeutic interventions. Exploring circRNA’s role in these applications may lead to significant advancements in disease treatment, immunotherapy, vaccine development, and diagnostic approaches.
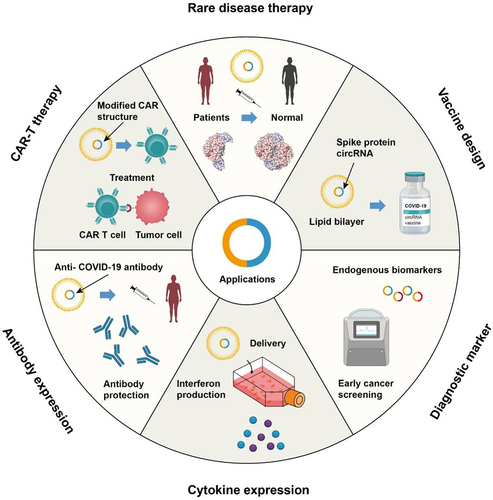
Figure 3. circRNA translating processes.
A. Cap-independent translation of circRNAs mediated by Internal Ribosome Entry Sites (IRES). IRES elements, featuring specialized secondary stem-loop structures located in the untranslated regions (UTRs) of circRNAs, directly engage and bind ribosomes, instigating translation through the formation of ribosome-IRES complexes. In circRNA, these recruitment proteins (eIF4G, eIF4A, eIF4B, and eIF3) are instrumental in facilitating the translation process. They assist in the initiation and regulation of protein synthesis, ensuring efficient progression. Specifically, eIF4G aids in the formation of the translation initiation complex, eIF4A is involved in unwinding mRNA structures, eIF4B enhances eIF4A’s unwinding activity, and eIF3 coordinates the assembly of the pre-initiation complex. This coordinated action of recruitment proteins is crucial for effective translation of circRNA.
B. Translation initiation in eukaryotic cells is triggered by the methylation of the sixth nitrogen in adenosine (m6A modification) within the UTR preceding the AUG triplets of circRNAs. This m6A modification is dynamically regulated through post-transcriptional processes involving ‘writers’ (methyltransferases), ‘erasers’ (demethylases), and ‘readers’ (proteins containing conserved m6A-binding domains). The reader protein YTHDF3, belonging to the YTH domain family, identifies m6A-circRNAs and recruits translation initiation factors like eukaryotic initiation factor 4 gamma 2 (eIF4G2) to initiate the translation of circRNAs.
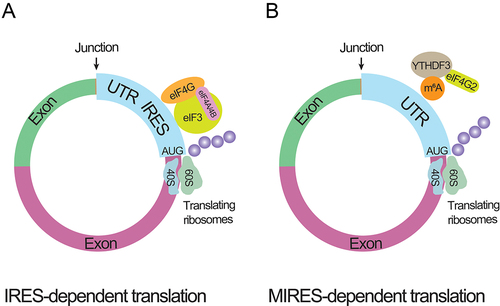
Table 2. The translational usage of circRNA.
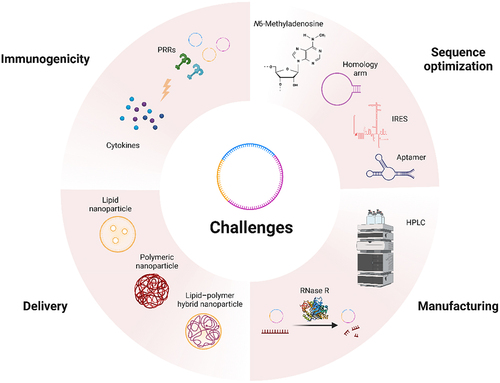
Figure 4. Challenges of circRNA nanomedicine.
Optimization of homologous arms is key to enhancing circulation efficiency of circRNA. The achievement of efficient translation is dependent on several factors, including adenosine modification (N6-methyladenosine), incorporation of IRES, and utilization of aptamers. Despite significant production advancements in recent years, purification and delivery of circRNA remain challenging. The immunogenic properties of circRNA require further investigation and exploration.
Data availability statement
Data sharing is not applicable to this article as no new data were created or analysed in this study.