Figures & data
Figure 1. Relationship between Arctic fox harvest and reproductive success (proportion of dens that produced a litter). Harvest data provided by Manitoba Wildlife and Fisheries Branch.
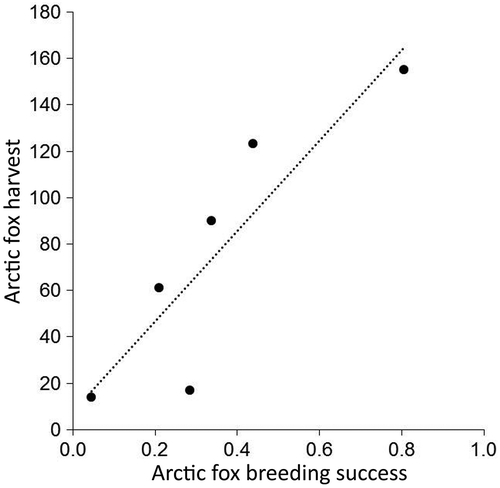
Figure 2. Relationship between Arctic fox reproductive success (proportion of dens that produced a litter) and density of (a) juvenile Canada geese (km‒2) and (b) rodents (log-transformed ha–1) the same year near Cape Churchill, Manitoba. Reproductive success was unrelated to juvenile goose density, but was positively related to rodent density (R2 = 0.718). Arctic fox breeding success reflects the proportion of dens that produced pups. Fox and rodent data for 1994‒97 were from Roth (Citation2002, Citation2003).
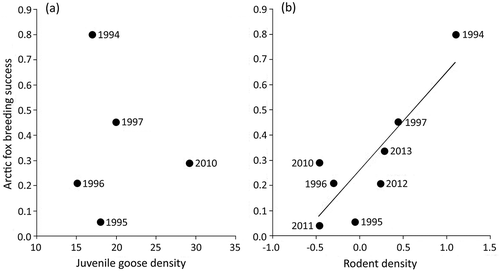
Table 1. The stable isotope ratios (mean ± SE) of Arctic fox hair grown in fall 1993–96 and 2009–2011. Carbon values from 1993–96 were adjusted to reflect atmospheric carbon in 2010 (Long et al. Citation2005). Years connected by the same letter did not differ (Tukey–Kramer HSD).
Table 2. Stable isotope ratios (mean ± SE) and mean C:N concentrations in samples of Arctic fox prey (eggs or muscle tissue) collected near Cape Churchill, Manitoba, Canada. Signatures varied among sources (MANOVA, Pillai = 1.534, F12,70 = 19.183, p < 0.001). Sources connected by the same letter had similar stable isotope ratios (p > 0.05), based on post-hoc tests for δ13C and δ15N that included ANOVA and Tukey’s HSD.
Table 3. The most likely relative contribution (mode) of prey sources to the Arctic fox fall diet estimated by the stable isotope-mixing model SIAR (Parnell et al. Citation2010). Values in parenthesis represent 90% credible intervals.
Figure 3. Mean (±SD) stable isotope ratios of Arctic fox winter hair (open circles) and prey samples (filled diamonds) in northern Manitoba. Fox hair values were corrected for trophic discrimination (Lecomte et al. Citation2011). Samples from 1994–97 (Roth Citation2002) were adjusted for the temporal change in atmospheric carbon to reflect projected values in 2010 (Long et al. Citation2005).
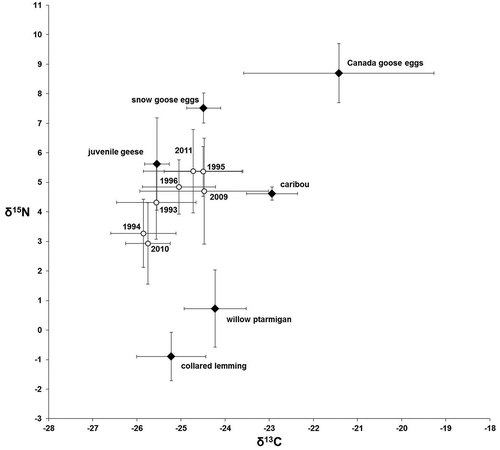
Figure 4. The proportion of juvenile geese in the Arctic fox diet was affected by that year’s density of (a) juvenile geese (km‒1; R2 = 0.620), but not (b) lemmings (log-transformed ha‒1). Diet contributions reflect the modal diet estimate from SIAR output.
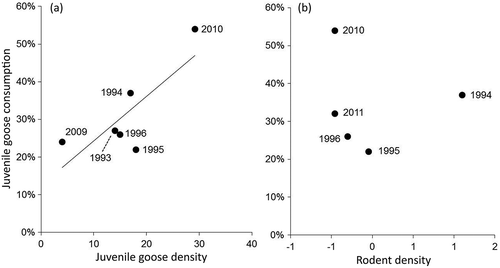
Figure 5. Relationship between rodent consumption and goose egg consumption by Arctic foxes in the fall of 1993–96 and 2009–2011 in northern Manitoba (R2 = 0.907). Consumption estimates are the modal diet proportions determined by SIAR (Parnell et al. Citation2010). The estimated contributions of Canada goose eggs and snow goose eggs were added together to estimate total egg consumption.
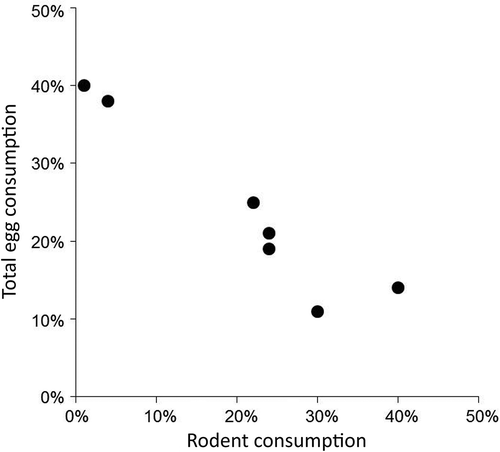
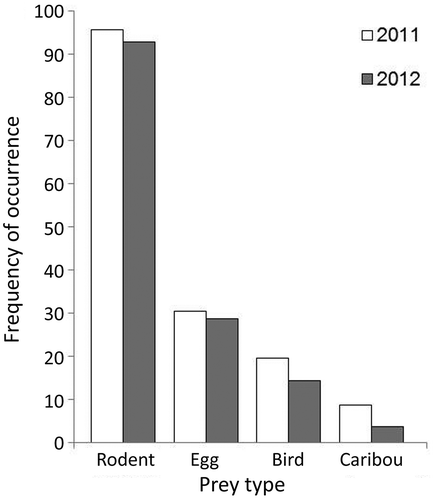
Figure 6. Winter diet of Arctic foxes in northern Manitoba estimated from scat samples collected from dens in April 2011 (n = 46) and 2012 (n = 28). Frequency of occurrence reflects the frequency of each prey type calculated by dividing the number of occurrences of a prey type by the number of faecal samples analysed in each year.