Figures & data
Table 1. Compositions of the different protein sources.
Table 2. Composition of the high fat high sucrose diets and the high fat high protein diets with different protein sources.
Table 3. Fatty acid compositions of the high fat high protein diets with different protein sources.
Figure 1. Effect of high fat high protein diets with different protein sources on body mass, body composition and digestibility. Male C57BL/6J mice were fed a high fat diet with a low protein:carbohydrate ratio (HF/HS) using casein as the protein source or high fat diets with a high protein:carbohydrate ratio (HF/HP) using different protein sources for 11 weeks. Mice fed a low fat regular diet (RD) were used as a reference. (A), Body mass development was recorded during the first 8 weeks. B-C , After 11 weeks epididymal white adipose tissue (eWAT) (B) and inguinal white adipose tissue (iWAT) (C) were dissected out and their masses recorded. D-E, Energy intake (D) and feed efficiency (E) were calculated based on data collected during the first 8 weeks of feeding. F-G, Apparent fat (F) and nitrogen (G) digestibility were calculated based on feed intake and feces collection during the 7th week of feeding. Data represent mean ± SEM (n = 9). Different small letters denote significant differences between the groups (p < 0 .05).
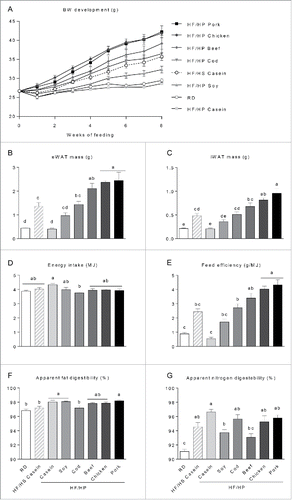
Figure 2. Effect of high fat diets with high and low protein:carbohydrate ratios on body mass gain, body composition and digestibility. Male C57BL/6J mice were fed a high fat diet with a low protein:carbohydrate ratio (HF/HS) or a high protein:carbohydrate ratio (HF/HP) using casein or pork as protein sources for 11 weeks. The mice were feed-deprived 4 h before blood collection and termination. (A), Body mass development was recorded and body weight (BW) gain calculated during the first 8 weeks of the trial. B-C, Fat mass (B) and lean mass (C) were measured after 8 weeks of feeding. D, Feed efficiency was calculated based on data collected during the first 8 weeks of feeding. E-F, Apparent fat (E) and nitrogen (F) digestibility were calculated based on feed intake and feces collection during the 7th week of feeding. G-I, Plasma levels of urea (G), hydroxybutyrate (H) and free fatty acids (FFAs) (I) were measured in plasma collected at termination. Data represent mean ± SEM (n = 9). Significant differences (p < 0 .05) between the protein sources are presented with different letters and differences between high and low protein:carbohydrate ratio with*.
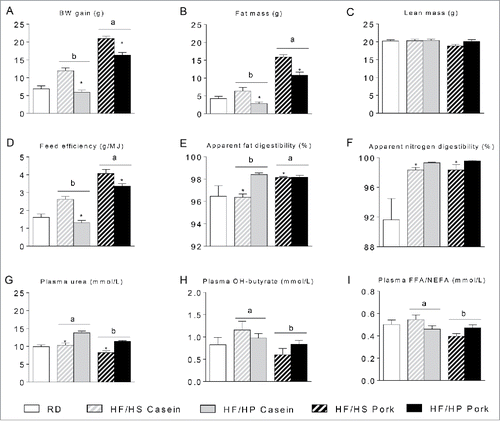
Table 4. Plasma concentrations of amino acids in mice fed high fat high sucrose (HF/HS) or high fat high protein (HF/HP) diets with casein or pork as the protein source.
Figure 3. Effect of high fat diets with high and low protein:carbohydrate ratios on activity, energy expenditure (EE) and respiratory exchange ratio (RER). Male C57BL/6J mice were fed high fat diets with a low protein:carbohydrate ratio (HF/HS) or a high protein:carbohydrate ratio (HF/HP) using casein or pork as protein sources. 48 hours measurements of activity (A) with total number of beam breaks (B). PRCF analysis of EE (C) and RER (F) over 48 hours with graphs illustrating the mean value. Based on the PRCF data EC50 (D,G) and hill slope (E,H) were calculated. Data represent mean ± SEM (n = 4). Significant difference (p < 0 .05) between the protein sources are presented with different letters and differences between high and low protein:carbohydrate ratio with *.
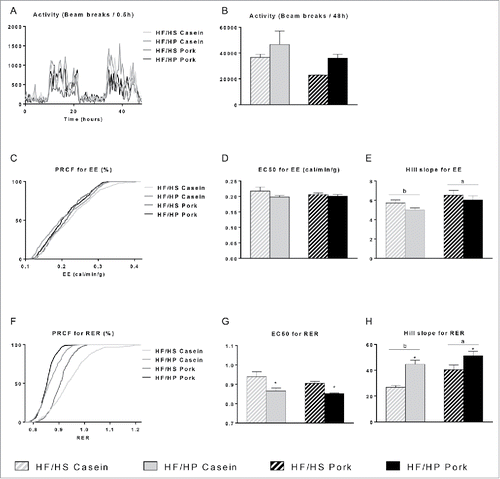
Figure 4. Effect of high fat diets with high and low protein:carbohydrate ratios on histological appearance and gene expression in inguinal white adipose tissue (iWAT). Male C57BL/6J mice were fed a high fat diet with a low protein:carbohydrate ratio (HF/HS) or a high protein:carbohydrate ratio (HF/HP) using casein or pork as protein sources for 11 weeks. The mice were feed-deprived 4h before termination. (A) Sections of iWAT (n = 4) were stained with eosin and hematoxylin, scalebar = 100 μm and (B) mean diameter of the adipocytes was measured and data represent mean ± SEM. (C) Expression levels of uncoupling protein 1 (Ucp1), deiodinase, iodothyronine, type II (Dio2), peroxisome proliferator-activated receptor-γ coactivator 1α (Ppargc1a) and cell death-inducing DFFA-like effector (Cidea) were measured and expression levels were normalized to TATA-box binding protein (Tbp). Gene-expression data represent mean ± SEM (n = 9). Significant difference (p < 0 .05) between the protein sources are presented with different letters and differences between high and low protein:carbohydrate ratio with *.
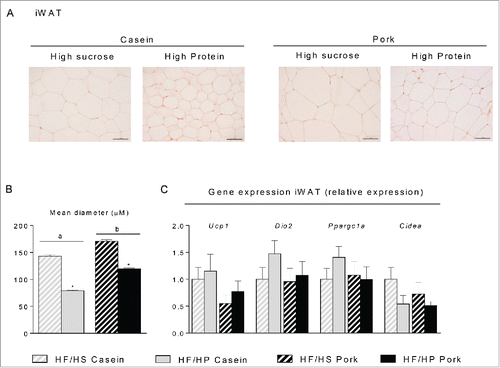
Figure 5. Effect of high fat diets with high and low protein:carbohydrate ratios on histological appearance, protein and gene expression in interscapular brown adipose tissue (iBAT). Male C57BL/6J mice were fed a high fat diet with a low protein:carbohydrate ratio (HF/HS) or a high protein:carbohydrate ratio (HF/HP) using casein or pork as protein sources for 11 weeks. The mice were feed-deprived 4 h before termination. (A) Sections of iBAT (n = 4) were stained with eosin and hematoxylin (H&E), scalebar = 100 μm and (B) Sections of iBAT (n = 4) were immunohistologically stained with an UCP1-antibody and (C) the area quantified, scalebar = 200μm. (D) UCP1 was detected by Western-blotting (n = 6) and the expression levels were quantified. (E) Expressions of uncoupling protein 1 (Ucp1) and Leptin were measured in addition to genes involved in fatty acid oxidation, carnitine palmitoyltransferase 1b (Cpt1b), acyl-Coenzyme A dehydrogenase, medium chain (Acadm) and acyl-Coenzyme A oxidase 1 (Acox1), fatty acid synthesis, fatty acid synthase (Fasn), stearoyl-Coenzyme A desaturase 1 (Scd1) and elongation of very long chain fatty acids (Elovl3) and triacylglycerol synthesis- and breakdown, microsomal CoA:glycerol-3-phosphate acyltransferase 4 (Gpat4), diacylglycerol acyltransferase 1 (Dgat 1), glycerol kinase (Gyk) glycerol-3-phosphate dehydrogenase 1 (Gpd1) and nuclear receptor interacting protein 1 (Nrip1). Expression levels were normalized to TATA-box binding protein (Tbp). Gene expression data represent mean ± SEM (n = 9). Significant difference (p < 0 .05) between the protein sources are presented with different letters and differences between high and low protein:carbohydrate ratio with *.
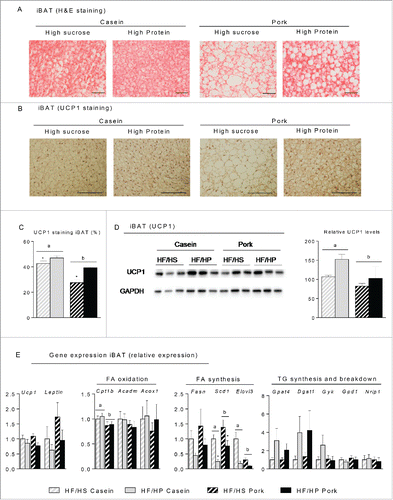
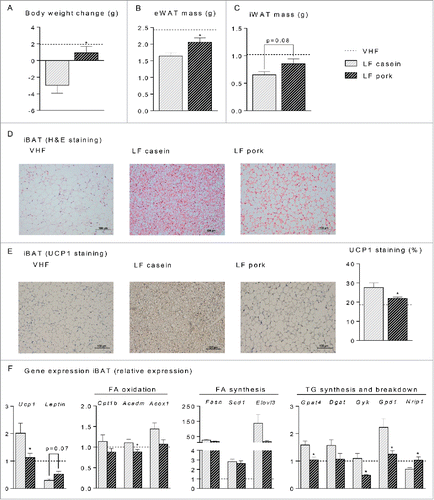
Figure 6. Effect of low fat (LF) diets with casein or pork as protein source on body and fat mass reduction, histological appearance and gene expression in interscapular brown adipose tissue (iBAT). Male C57BL/6J mice were fed a very high fat (VHF) diet for 11 weeks to induce obesity development and were then switched to low fat (LF) diets (n = 10) containing either casein or pork as protein source for 6 additional weeks. One group marked with a scattered line continued eating the VHF diet. (A) body weight change of the mice after 6 weeks on the LF diets. The mice were terminated and (B) the epididymal white adipose tissue (eWAT) mass and (C) the inguinal white adipose tissue (iWAT) mass were recorded. (D) sections of iBAT (n = 4) were stained with eosin and hematoxylin (H&E), scalebar = 100 μm (E) sections of iBAT (n = 4) were immunohistologically stained with an UCP1-antibody and the area quantified, scalebar = 200 μm. (F) expression level of uncoupling protein 1 (Ucp1) and Leptin in addition to genes involved in fatty acid oxidation, carnitine palmitoyltransferase 1b (Cpt1b), acyl-Coenzyme A dehydrogenase, medium chain (Acadm) and acyl-Coenzyme A oxidase 1 (Acox1), fatty acid synthesis, fatty acid synthase (Fasn), stearoyl-Coenzyme A desaturase 1 (Scd1) and elongation of very long chain fatty acids (Elovl3) and triacylglycerol synthesis- and breakdown, microsomal CoA:glycerol-3-phosphate acyltransferase 4 (Gpat4), diacylglycerol acyltransferase 1 (Dgat 1), glycerol kinase (Gyk) glycerol-3-phosphate dehydrogenase 1 (Gpd1) and nuclear receptor interacting protein 1 (Nrip1). Expression levels were normalized to TATA-box binding protein (Tbp). Expression of individual genes in mice fed the LF diet is expressed relative to the expression in the mice fed the VHF (scattered line) diet. Significant differences (p < 0 .05) between the protein sources are presented with*.