Figures & data
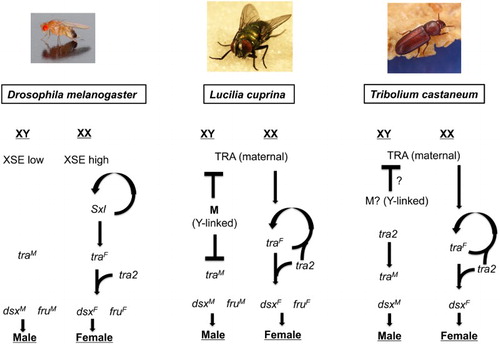
Figure 1. A conditional Cas9-mediated gene drive. The tetracycline transactivator (tTA) is expressed in the germline using the vasa gene promoter. In the absence of tetracycline in the diet, tTA will bind to tetO and activate expression of Cas9. Expression of a gRNA complementary to an essential gene is controlled using a U6 gene promoter. Following Cas9 digestion of the essential gene, homology-directed repair will lead to insertion of the multi-gene cassette bracketed by left (LHA) and right (RHA) homology arms. A GFP marker gene would facilitate initial identification of genetically modified insects. Insulator elements (I) prevent tTA bound to tetO from enhancing its expression from the vasa gene promoter, which could reduce fitness. Additional containment measures should be used for drive evaluation such as physical barriers and splitting the drive by not including the vasa-Cas9 or U6-gRNA gene in the construct shown.
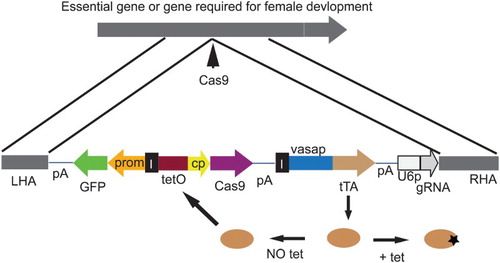
Figure 2. A comparison of the genetic mechanisms that determine sex in Drosophila melanogaster, Lucilia cuprina and Tribolium casteneum. In female D. melanogaster embryos, a high dose of X signal elements (XSE) leads to activation of the master gene Sex lethal (Sxl). Sxl both autoregulates its expression and switches transformer (tra) RNA splicing into the female mode. Only the female form of tra mRNA codes for functional protein. TRA combines with TRA-2 to switch doublesex (dsx) and fruitless (fru) into the female modes of splicing. DSX and FRU control the expression of many genes that influence sexual development and behavior. The bottom of the regulatory hierarchy in L. cuprina appears to be the same as Drosophila. However, it is tra RNA splicing that is autoregulatory in females. The presence of the Y-linked M factor inhibits the female-mode of tra RNA splicing in male embryos. As in Drosophila, only the female form of tra mRNA codes for functional protein. In female embryos, maternal TRA likely contributes to the initiation of the female mode of tra splicing. The sex determination regulatory mechanisms appear to be identical in L. cuprina and C. hominivorax. Remarkably, the genetic mechanisms that determine sex in the red flour beetle, T. casteneum, appear to be very similar to the L. cuprina regulatory hierarchy. Adapted from Scott, Pimsler, and Tarone (Citation2014) and Shukla and Palli (Citation2013).