Figures & data
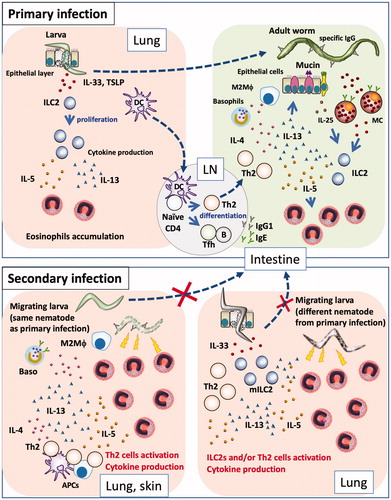
Figure 1. Primary and secondary immune responses in helminth infections. (Primary infection) Nematode infection induces differentiation of Th2 cells to induce Th2-type immune responses, such as mucus production and mastocytosis, as well as proliferation and accumulation of eosinophils and basophils, production of nematode antigen-specific antibodies, and differentiation of M2 macrophages. Some nematodes are expelled by these effectors, while others evade these effectors and develop chronic infection. Some nematodes exhibit a lung stage before they become adults in the intestines. When larvae penetrate the lung, they damage lung tissue, which leads to the release of epithelial cell-related cytokines (e.g., IL-33, TSLP, and IL-25) from epithelial and stromal cells; these cytokines induce proliferation of group 2 innate lymphoid cells and subsequent production of cytokines (e.g., IL-5 and IL-13), resulting in the accumulation of eosinophils. Thus, nematode infection induces pulmonary eosinophilia at 1–2 weeks after infection. However, eosinophils cannot damage larvae in the lungs, because nematodes have already moved to the intestines and developed into adult worms. (Secondary infection) If a host is re-infected by a species of nematode that it has previously encountered, Th2 cells and antibodies respond to the parasites robustly and promptly, thus trapping and damaging them at the sites of infection (e.g., skin) and at the lungs, through the actions of effector cells such as basophils and macrophages, prior to migration of the larvae to the intestines. If a host is infected by a species of nematode that it has not previously encountered, Th2 cells and antibodies cannot respond to the parasite antigen with effectiveness similar to that of the response to the sensitizing parasite; thus, the larvae can migrate to the lungs. Following tissue damage by migrating larva, stromal cells produce IL-33. The lungs of an infected host have large numbers of Th2 cells and/or ILC2s, which rapidly react to IL-33 by producing Th2 cytokines. Thus, infected hosts exhibit non-antigen-specific resistance by attacking migratory larvae through the actions of rapidly accumulated eosinophils.