Figures & data
Figure 1 Properties of low-level light. Sunlight is composed of a combination of noncoherent waves with wavelengths spanning the entire visible spectrum. In contrast, lasers emit waves of a single wavelength (monochromatic) that have spatial and temporal synchronization. This high wavelength coherence allows the transmission of energy at a high power density. Finally, low-level light consists of monochromatic or quasimonochromatic waves taking different paths leading to a common target point. While wavelength, radiant exposure, irradiance, and fractionation scheme are relevant for low-level light therapy applications, the authors introduce the possibility that noncoherence may be advantageous for some neurometabolic purposes. Noncoherence allows nervous tissue exposure at “therapeutic” wavelengths at relatively low power densities during the time necessary to modulate neural metabolism in response to activation or injury, even if this time is prolonged.
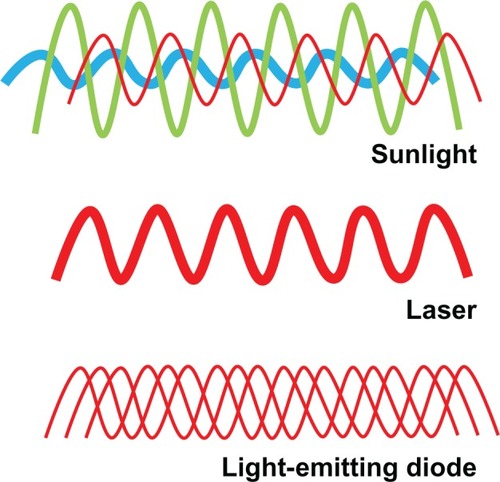
Figure 2 Principles of light-tissue interactions. (A) Light at short wavelengths has low tissue penetration. Light at high wavelengths displays high tissue penetration and delivers therapeutic levels of energy to deeper structures. Whereas surface structures exposed to high wavelengths may be exposed to inhibitory energy densities (eg, 100 J/cm2), light with a certain power density targeting a surface redistributes in a proportionally higher tissue volume due to diffraction (bending of waves). Multiple scattering of light allows for spreading out of waves and increases the treatment volume, so a lower applied energy can be used to achieve an effective energy density at higher depth. (B) Because radiant exposure (J/cm2) is the product of irradiance (W/cm2) and time, the energy delivered to tissues as a result of a constant irradiance can be increased by increasing exposure time. Thus, tissue penetration can also be affected by exposure time. When sources of low-level light therapy are used with high exposure times, deep structures can be treated with biomodulatory amounts of energy, while avoiding ablative effects. (C) Finally, tissues vary in their photoacceptor content, transmittance, and relaxation time. This accounts for interspecies and interregional variations in light penetration (eg, gray matter versus white matter in the brain). In addition, metabolically active tissues such as nervous tissue may exhibit variations in relaxation times, due to changes in the redox states of photoacceptors. This not only potentially affects tissue penetration, but also the susceptibility of nervous tissues to low-level light therapy depending on their activational state.
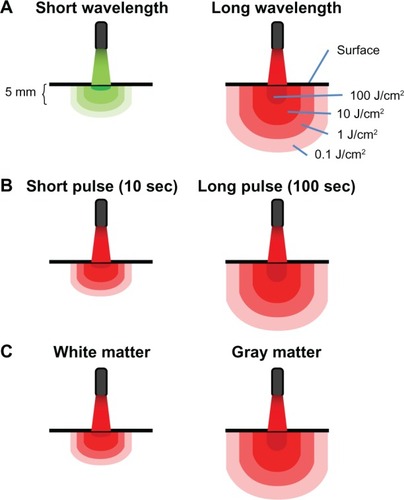
Table 1 Major parameters of low-level light therapy (LLLT)
Figure 3 Hormetic effects of low-level light therapy (LLLT). LLLT does not induce classical linear dose-response pharmacological effects. LLLT effects are characterized by inverted U-shaped dose-response curves, in which linear responses may be seen only at very low doses. Whereas linear effects may be negligible, maximal stimulatory effects are typically observed at intermediate doses. However, the linear relationship does not hold at high doses, since inhibitory effects are observed instead. In fact, the inhibitory effects of very high LLLT doses might be worse than control conditions (eg, tissue destruction). A key observation concerning the modulatory effects of light in tissues is that maximal responses at intermediate doses tend to represent less than twofold increases in biological variables relative to baseline conditions. Yet these effects have been shown to have major relevance, especially when energy metabolism is involved in nervous tissue. Thus, hormesis is an essential concept for the development of neurotherapeutic applications of LLLT.
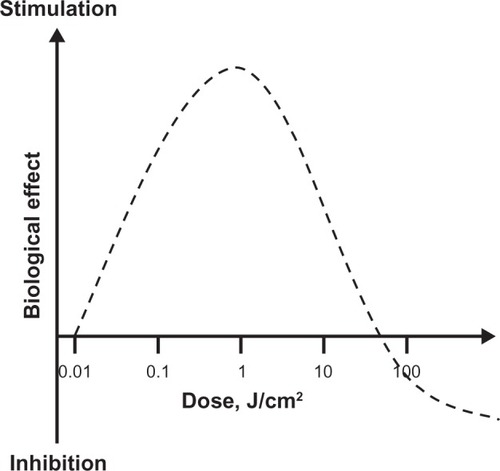
Figure 4 Differential effects of light on photoreceptors and photoacceptors in the retina. Light reaches the retina and travels through the different retinal layers to reach the outermost photoreceptor layer. It then excites the photoreceptor rhodopsin in rods and cones, triggering the process of phototransduction. Phototransduction causes photoreceptor cell hyperpolarization, changes in neurotransmission, and action potentials (yellow arrows) in bipolar cells and ganglion cells. These effects represent the onset of visual information processing. Light can also directly excite photoacceptors in neurons including retinal ganglion cells. The main photoacceptor in the red to near-infrared spectrum is the mitochondrial respiratory enzyme cytochrome oxidase. The effects of light on neuronal cytochrome oxidase induce modulation of cell bioenergetic mechanisms that are independent from visual processing. Yet photobiomodulation has major implications in neuronal physiology and homeostasis.
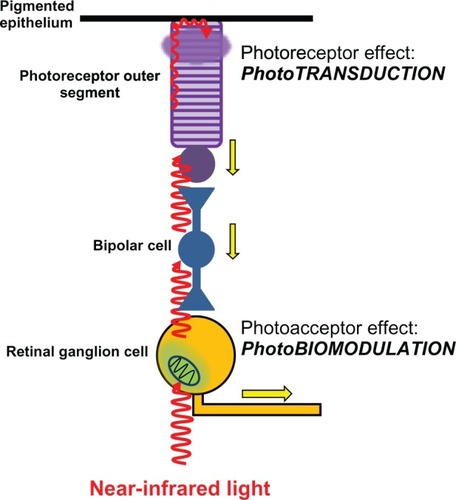
Figure 5 The intracellular mechanisms of action of low-level light therapy (LLLT). Photobiomodulation results in a cascade of intracellular pleiotropic effects. Light is absorbed by chromophores in cytochrome oxidase and induces changes in its redox state. Redox reaction of enzymes in the inner mitochondrial membrane induces accelerated electron flow, reduced nicotine adenine dinucleotide (NADH) consumption and increase in the mitochondrial membrane potential. These changes facilitate the synthesis of adenosine triphosphate (ATP) and increase the generation of free radicals. Increased ATP availability allows the activation of kinases that induce the release of calcium and the formation of cyclic adenosine monophosphate (cAMP). Calcium, cAMP, and free radicals act as second messengers and are able to activate different metabolic pathways at the nuclear level. Depending on the cell environment, these cellular changes can be adaptive and promote enhancement of neuronal physiology that translates in clinical improvement.
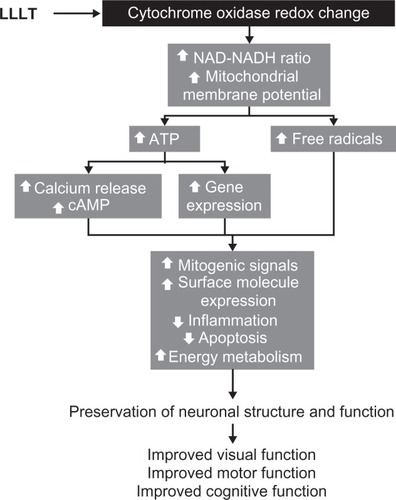
Table 2 Beneficial in vivo effects of low-level light therapy on the eye
Table 3 Beneficial in vivo transcranial effects of low-level light therapy on the brain