New insights on the role of epigenetic alterations in hepatocellular carcinoma
Maddalena Frau1 Department of Clinical and Experimental Medicine, Division of Experimental Pathology and Oncology, University of Sassari, Sassari, Italy
,
Claudio F Feo2 Department of Clinical and Experimental Medicine, Division of Surgery, University of Sassari, Sassari, Italy
,
Francesco Feo1 Department of Clinical and Experimental Medicine, Division of Experimental Pathology and Oncology, University of Sassari, Sassari, ItalyCorrespondence[email protected]
&
Rosa M Pascale1 Department of Clinical and Experimental Medicine, Division of Experimental Pathology and Oncology, University of Sassari, Sassari, Italy
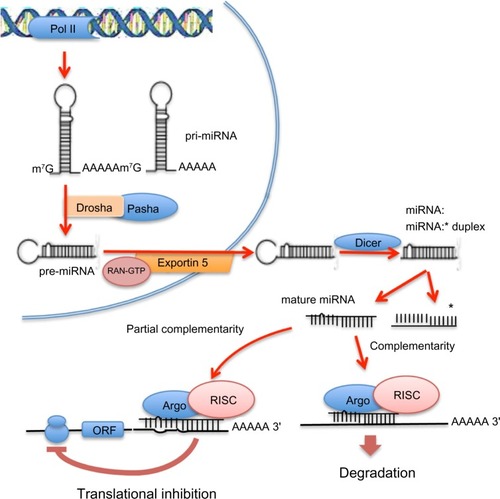
Figure 1 Intranuclear and extranuclear phases of the process of maturation of microRNAs (miRNAs) and mechanisms of inhibition of messenger RNA transcription. In the nucleus, long primary transcripts (primary miRNAs) are cleaved by RNase III Drosha, with its cofactor Pasha, to release the precursor miRNAs, which are exported to the cytoplasm by exportin 5. In the cytoplasm, precursor miRNAs are processed by RNase III Dicer, with the release double-stranded duplexes of 20–23 RNA that contain the mature miRNA and the passenger miRNA strand (asterisked). The mature miRNAs interact with the 3′ untranslated region of its targeted mRNAs to form, with Argonaute proteins, the RNA-induced silencing complex (RISC). RISC binds to the 3′ untranslated region of its target or targets, inhibiting translation of single or multiple proteins; complete miRNA complementarity induces mRNA degradation, whereas partial complementarity represses translation.
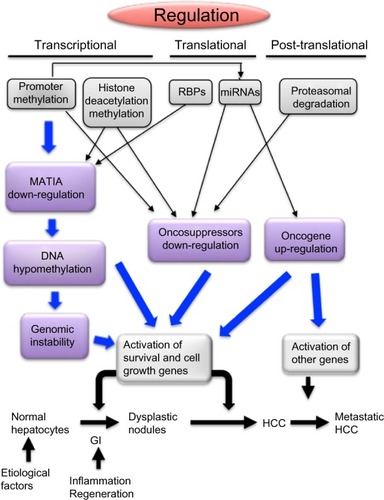
Figure 3 Transcriptional, translational, and posttranslational mechanisms leading to deregulation of MAT1A, tumor suppressor genes, and oncogenes during hepatocarcinogenesis.
LinJHuangSWuSMicroRNA-423 promotes cell growth and regulates G(1)/S transition by targeting p21Cip1/Waf1 in hepatocellular carcinomaCarcinogenesis201132111641164721890460
YangLMaZWangDZhaoWChenLWangGMicroRNA-602 regulating tumor suppressive gene RASSF1A is overexpressed in hepatitis B virus-infected liver and hepatocellular carcinomaCancer Biol Ther201091080380820364114
WongQWChingAKChanAWMiR-222 overexpression confers cell migratory advantages in hepatocellular carcinoma through enhancing AKT signalingClin Cancer Res201016386787520103675
GarofaloMDi LevaGRomanoGmiR-221&222 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 downregulationCancer Cell200916649850919962668
GramantieriLFornariFFerracinMMicroRNA-221 targets Bmf in hepatocellular carcinoma and correlates with tumor multifocalityClin Cancer Res200915165073508119671867
XieQHHeXXChangYMiR-192 inhibits nucleotide excision repair by targeting ERCC3 and ERCC4 in HepG2.2.15 cellsBiochem Biophys Res Commun2011410344044521672525
XuTZhuYXiongYGeYYYunJPZhuangSMMicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of human hepatocellular carcinoma cellsHepatology200950111312119441017
GramantieriLFerracinMFornariFCyclin G1 is a target of miR-122a, a microRNA frequently down-regulated in human hepatocellular carcinomaCancer Res200767136092609917616664
WongQWLungRWLawPTMicroRNA-223 is commonly repressed in hepatocellular carcinoma and potentiates expression of Stathmin1Gastroenterology2008135125726918555017
ZhangJPZengCXuLGongJFangJHZhuangSMMicroRNA-148a suppresses the epithelial-mesenchymal transition and metastasis of hepatoma cells by targeting Met/Snail signalingOncogene Epub992013
SalviASabelliCMonciniSMicroRNA-23b mediates urokinase and c-met downmodulation and a decreased migration of human hepatocellular carcinoma cellsFEBS J2009276112966298219490101
WongCCWongCMTungEKThe microRNA miR-139 suppresses metastasis and progression of hepatocellular carcinoma by down-regulating Rho-kinase 2Gastroenterology2011140132233120951699
FanQHeMDengXDerepression of c-Fos caused by microRNA-139 down-regulation contributes to the metastasis of human hepatocellular carcinomaCell Biochem Funct201331431932423001723
LiDLiuXLinLMicroRNA-99a inhibits hepatocellular carcinoma growth and correlates with prognosis of patients with hepatocellular carcinomaJ Biol Chem201128642366773668521878637
FanCGWangCMTianCmiR-122 inhibits viral replication and cell proliferation in hepatitis B virus-related hepatocellular carcinoma and targets NDRG3Oncol Rep20112651281128621725618
HeXXChangYMengFYMicroRNA-375 targets AEG-1 in hepatocellular carcinoma and suppresses liver cancer cell growth in vitro and in vivoOncogene201231283357336922056881
LiuAMPoonRTLukJMMicroRNA-375 targets Hippo-signaling effector YAP in liver cancer and inhibits tumor propertiesBiochem Biophys Res Commun2010394362362720226166
LiNFuHTieYmiR-34a inhibits migration and invasion by down-regulation of c-Met expression in human hepatocellular carcinoma cellsCancer Lett20092751445319006648
LuYYueXCuiYZhangJWangKMicroRNA-124 suppresses growth of human hepatocellular carcinoma by targeting STAT3Biochem Biophys Res Commun2013441487387924211205
ZhangJFHeMLFuWMPrimate-specific microRNA-637 inhibits tumorigenesis in hepatocellular carcinoma by disrupting signal transducer and activator of transcription 3 signalingHepatology20115462137214821809363
YoonSKimTHNatarajanAAcute liver injury upregulates microRNA-491-5p in mice, and its overexpression sensitizes Hep G2 cells for tumour necrosis factor-alpha-induced apoptosisLiver Int201030337638720015148
TsaiWCHsuPWLaiTCMicroRNA-122, a tumor suppressor microRNA that regulates intrahepatic metastasis of hepatocellular carcinomaHepatology20094951571158219296470
ZhangJYangYYangTmicroRNA-22, downregulated in hepatocellular carcinoma and correlated with prognosis, suppresses cell proliferation and tumourigenicityBr J Cancer201010381215122020842113
TakataAOtsukaMKojimaKMicroRNA-22 and microRNA-140 suppress NF-κB activity by regulating the expression of NF-κB coactivatorsBiochem Biophys Res Commun2011411482683121798241
LinCJGongHYTsengHCWangWLWuJLmiR-122 targets an anti-apoptotic gene, Bcl-w, in human hepatocellular carcinoma cell linesBiochem Biophys Res Commun2008375331532018692484
WeiXXiangTRenGmiR-101 is down-regulated by the hepatitis B virus x protein and induces aberrant DNA methylation by targeting DNA methyltransferase 3ACell Signal201325243944623124077
WangCMWangYFanCGmiR-29c targets TNFAIP3, inhibits cell proliferation and induces apoptosis in hepatitis B virus-related hepatocellular carcinomaBiochem Biophys Res Commun2011411358659221763284
HuangSHeXDingJUpregulation of miR-23a approximately 27a approximately 24 decreases transforming growth factor-beta- induced tumor-suppressive activities in human hepatocellular carcinoma cellsInt J Cancer2008123497297818508316