Figures & data
Figure 1. Architecture of the adult Drosophila visual system. (A) The optic lobe consists of the lamina, medulla, lobula plate, and lobula. The illustration shows a subset of neuron subtypes and their retinotopic projections. R1–R6 cells extend axons from the retina into the lamina, where they connect with lamina neurons L1–L3 in cartridges. Axons of R8 and R7 cells and lamina neurons L1–L5 terminate in one or more of ten medulla neuropil layers (M1–M10). Transmedullary neurons (Tm and TmY) project from the medulla to subsets of six lobula (Lo) and four lobula plate (Lop) neuropil layers. Distal medulla (Dm) neurons innervate several columns in upper medulla layers. Medulla intrinsic (Mi) neurons connect distal and proximal layers. C neurons extend branches into the medulla and lamina. T neurons connect the lobula or lobula plate and medulla, or lobula and lobula plate. (B) R1–R6 axons innervate lamina cartridges following the neural superposition principle.
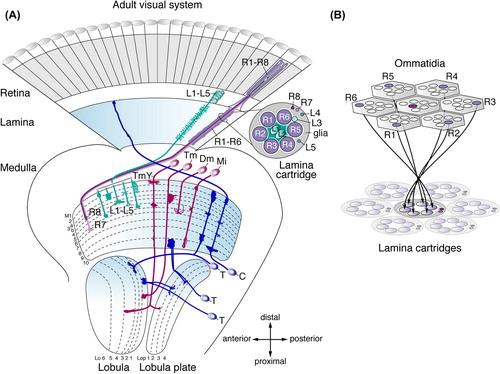
Figure 2. Early optic lobe development and lamina neurogenesis. (A) Embryonic stages 11 and 12: the optic placode forms in the posterior procephalic region of the head ectoderm, invaginates (arrowhead), and attaches to the central brain. Ventral cells develop into Bolwig's organ (BO), containing the larval photoreceptor cells. At the early first instar larval stage, the outer and inner proliferation centers (OPC and IPC) are visible. They expand by symmetric divisions and become crescent-shaped during the late first and early second instar larval stages, respectively. The OPC and IPC separate, as they begin to produce offspring from the early third instar larval stage onward. (B) In the third instar larval eye imaginal disc, rows of R-cell clusters progressively arise posteriorly (p) of the morphogenetic furrow (MF). R8 cells sequentially recruit R2/R5, R3/R4, R1/R6, and R7 cells into each cluster. A lateral view of the optic lobe shows the crescents of the lamina, the OPC neuroepithelium (NE) and medulla NBs, GMCs, and medulla neurons (mn). Lamina neurogenesis occurs in the wake of the lamina furrow (LF). Glial precursor cell (GPC) areas are located at the dorsal and ventral tips of the OPC crescent. (C) A horizontal view of the third instar larval eye imaginal disc and optic lobe shows the stepwise formation of lamina neurons (ln) and medulla neurons. Medially, the OPC NE gives rise to medulla NBs, GMCs and medulla neurons. Laterally, the OPC NE generates lamina precursor cells (LPCs). The position of the NE to LPC conversion at the bottom of the lamina furrow is approximate in this schematic presentation. LPCs give rise to undifferentiated lamina neurons in the pre-assembly domain (pAD). Lamina neurons assemble into columns in close association with R-cell bundles and differentiate into neuronal subtypes L1–L5. The IPC produces neurons of the lobula complex. (D) R-cell axons provide the anterograde signals Hedgehog (Hh) and Spitz (sSpi) to control LPC proliferation, lamina column assembly and lamina neuron differentiation. a, anterior; Bsh, Brain-specific homeobox; Ci, Cubitus interruptus; Dac, Dachshund; EGFR, epidermal growth factor receptor; Elav, Embryonic lethal abnormal vision; Hbs, Hibris; Sim, Single-minded; Rst, Roughest.
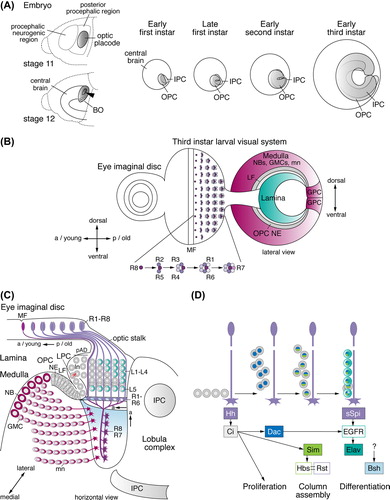
Figure 3. Medulla neurogenesis depends on the sequential conversion of neuroepithelial (NE) cells to NBs. (A) Outer proliferation center (OPC) NE cells gradually convert into medulla NBs in a medial to lateral orientation. The advancement of the proneural wave is defined by Lethal of scute (L'sc) expression. (B) The gradual conversion of NE cells to NBs involves a switch from symmetric to asymmetric cell divisions. The transition zone consists of progenitors PII expressing L'sc and PI expressing low levels of Deadpan (Dpn). NBs express Dpn and Asense (Ase). GMCs express Ase and Prospero (Pros). Newly formed medulla neurons (mn) maintain Pros. (C) The progression of the proneural wave from medial to lateral is negatively regulated by the Notch (N) pathway and positively regulated by the epidermal growth factor receptor (EGFR) pathway. N signaling is activated in PI NE cells by Delta (Dl) in PII and by Serrate (Ser) in surface glia (sg). Dl expression is repressed by Ecdysone (20-HE, 20-Hydroxyecdysone) signaling. EGFR signaling activates Pointed P1 (PntP1) and Rhomboid (Rho), which promote the secretion of Spitz (sSpi) from PII. sSpi is also provided by a cortex glia subtype (cg) expressing the microRNA miR-8. N and EGFR signaling regulate each other. The JAK/STAT and Fat/Hippo signaling pathways provide additional negative and positive inputs, respectively. LF, lamina furrow; LPC, lamina precursor cells.
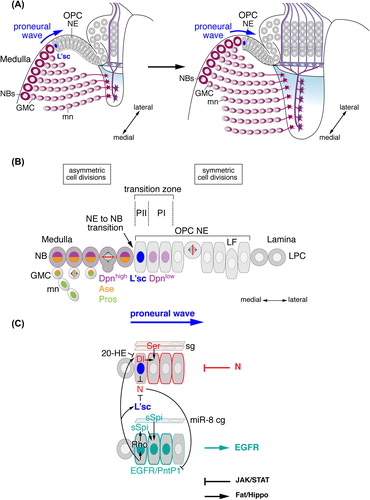
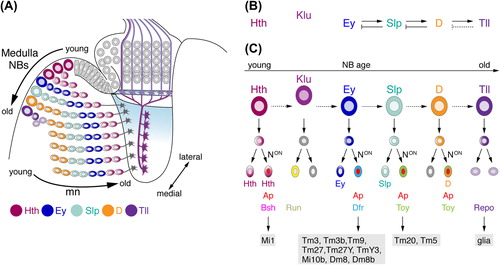
Figure 4. Temporal patterning of medulla NBs and Notch (N) signaling contribute to the generation of diverse neuron subtypes. (A) NE cells gradually convert into NBs. As medulla NBs age, they sequentially express the transcription factors Homothorax (Hth), Eyeless (Ey), Sloppy paired 1 and 2 (Slp), Dichaete (D), and Tailless (Tll). Each NB produces a column of medulla neurons. Progeny maintain the expression of the determinant present in the NB at the time of their birth. Older NBs and their offspring are located more medially. In each lineage, the oldest neurons are positioned closest to the medulla neuropil. Klumpfuss (Klu) is expressed in NBs but not in progeny and therefore has not been included in this schematic. Interim stages, during which NBs express more than one factor and neurons downregulating factors are also not shown. (B) Ey, Slp, and D are required for the transition to the next determinant. Slp, D, and Tll are necessary to repress the preceding factor in the series. Tll is sufficient but not required to repress D (dashed line). (C) N-mediated binary cell fate choices further diversify lineages. Progeny, in which N signaling is on, express Apterous (Ap). Depending on the combination of transcription factors present, the expression of subtype-identity determinants such as Brain-specific homeobox (Bsh), Runt (Run), Drifter (Dfr), or Twin of Eyeless (Toy) is induced. Lineages defined by the combinatorial expression of these determinants give rise to specific neuronal subtypes. Tll-positive NBs generate Reversed polarity (Repo)-positive glia. Dm, distal medulla neurons; Mi, medulla intrinsic neurons; Tm, transmedullary neurons; TmY, transmedullary Y neurons.