Figures & data
Fig. 1 Location of stations sampled for tintinnids. The green box distinguishes the polynya stations (following Lee et al. Citation2012).
Fig. 2 Water column profiles of (a, b) station 5, typical of deep off-shore stations, and (c, d) a polynya station, station 18. Note the difference in scales of the chlorophyll profiles. For further details see and 2.
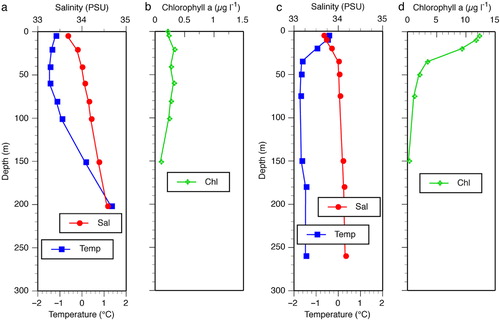
Fig. 3 Low magnification (4× objective) views of settled net tow material from (a) an offshore deep-water station, station 5, showing a dominance by diatoms, and (b) a coastal polynya station, station 18, showing a dominance by colonies of Phaeocystis. The area shown is equivalent to a volume of the original net tow material from about 1 L of seawater. The arrows show tintinnids (Cymatocylis).

Table 1 Locations and physical characteristics of the sample stations (see also ). Numbers in boldface denote polynya stations and one marginal polynya station (station 8), which was not completely free of sea ice. Station type, mixed layer and euphotic zone depth from, and methods described by Lee et al. (Citation2012).
Table 2 Biological characteristics of the stations. The primary production data (14C-based) are from in Lee et al. (Citation2012), estimated at a few of the stations sampled for microplankton. Microphytoplankton composition, based on examination of net tow material, was clearly dominated by either diatoms (D), Phaeocystis (P) or mixture of both (D/P). Tintinnid concentrations and number of species are shown for the top 150 m of the water column. Values in boldface denote data corresponding to polynya stations or near polynya conditions (station 8).
Fig. 4 Tintinnid species encountered in the Amundsen Sea samples. (a) Salpingella laackmanni, (b) Salpingella decurtata, (c) Salpingella faurei, (d) Laackmanniella naviculaefera, (e) Laackmanniella prolongata, (f) Amphorellopsis quinquelata, (g) Amphorides laackmanni, (h) Acanthostomella obtusa, (i) Codonellopsis pusilla, (j) Epiplocylcoides reticulata; (k) Codonellopsis gaussi, (l) Codonellopsis gaussi, (m) Codonellopsis gaussi forma globosa, (n) Codonellopsis gaussi forma cylindricoconica, (o) Condonellopsis gaussi forma coxiella, (p) Cymatocylis affinis/convallaria, (q) Cymatocylis affinis/convallaria forma calcyformis, (r) Cymatocylis affinis/convallaria forma subrotundata, (s) Cymatocylis affinis/convallaria forma drygalski and (t) Cymatocylis affinis/convallaria forma cylindrica. Note that some species agglutinate diatoms; the diatoms are not necessarily the most abundant according to Wasik et al. (Citation1996).

Table 3 Species records for each station with the station numbers in boldface denoting the polynya stations or near polynya conditions (station 8). The distributional type is based on the biogeographic classifications presented by Dolan et al. (Citation2012). Note that most species were absent from the polynya stations. The species found in polynya were exclusively the Southern Ocean endemic forms except for Salpingella faurei. Note that the species Codondellopsis gaussi and Cymatocylis convallaria group forms previously considered separate species in some reports.
Fig. 5 Scatterplot showing relationship between tintinnid and chlorophyll concentrations in the top 150 m. Symbols distinguish stations in which the microphytoplankton was dominated by diatoms, Phaeocystis, or was a mixture of both. There was no significant relationship between tintinnid and overall chlorophyll concentration or among stations in which diatoms dominated the microplankton community. While we have quantitative data for only three of the four polynya stations, the concentration of tintinnids does, however, appear to increase with chlorophyll concentrations in the Phaeocystis-dominated communities (n=3, r=0.987).
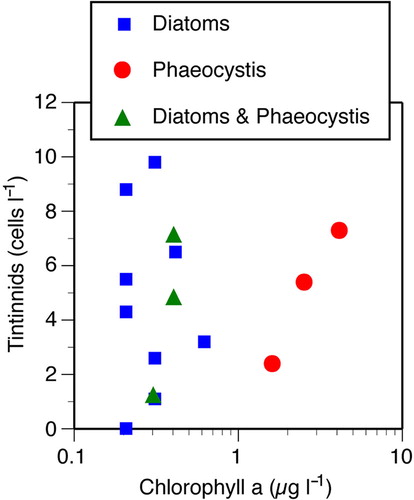
Fig. 6 Scatterplots showing the relationship between the numbers of different forms of (a) Cymatocylis affinis and (c) Condonellopsis gaussi found in relation to the concentration of chlorophyll (symbols show the dominant phytoplankton forms), and (b, d) the total concentrations of the species, all forms pooled. Note that there is no apparent relationship between numbers of forms and the concentration or composition of the phytoplankton. However, the number of forms found in a station sample was positively related to the overall concentration of Cymatocylis affinis, all forms pooled. For C. gaussi the relationship was not significant.
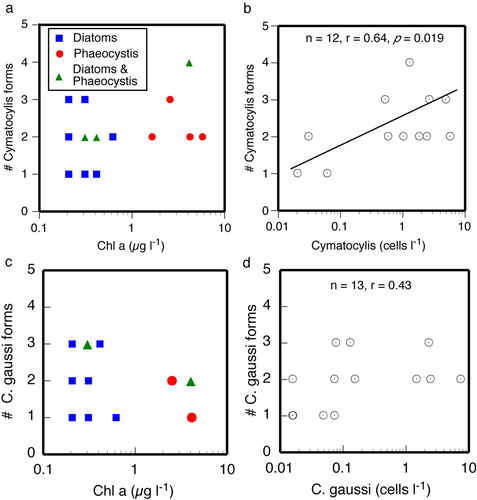
Table 4 Species records of the Amundsen Sea and the adjacent Bellinghausen Sea. Species names in boldface are endemic to the Southern Ocean; the others are known from a wide range of areas including other regions of the Southern Ocean (Dolan et al. Citation2012). Species now known to be forms or morphological variants of Cymatocylis affinis/convallaria and variants of Condonellopsis gaussi (see ) are not listed separately.