Figures & data
Figure 1 Hypothetical pre-karyote cell cycle and life cycle controlled by ancient DNA damage checkpoint. The haploid pre-karyote is assumed to start DNA replication (A). If there was no DNA damage, DNA replication would be completed (B) resulting in diploid entity (C), that would be allowed to divide to produce two haploid pre-karyotes (D). If there were DNA damage (E) resulting in broken replication forks, DNA damage checkpoint would stop replication to allow the DNA repair and the restart of DNA synthesis via recombiantion with the matrix DNA (F). After completing the DNA replication (G), diploid pre-karyote (C) would be allowed to divide and the result would be two haploid entities (D). If the DNA damage were huge and pre-karyote would not be able to repair it and restart the DNA synthesis by itself (H), it could fuse with the partner (perhaps damaged too) (I). After the repair of the damage and the restart of DNA synthesis via recombination with the DNA of the partner (J), replication of both haploid sets would result in a tetraploid entity (K). Now the cell division would be allowed (L) resulting in two diploid entities. However, these would have no need to start replication again, instead both would be allowed to divide again (M and C) and the result would be four haploid entities (N and D).
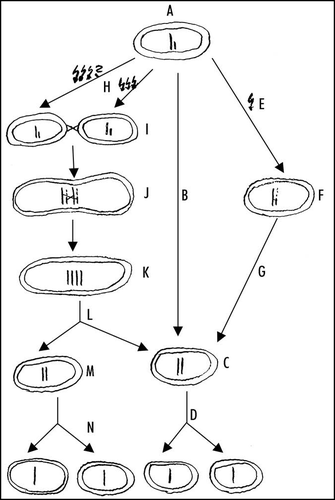
Figure 2 The hypothetical scenarios for the origin of two-membrane-bounded LUCA. (A) The fusion occurring at the orifice of a single gastruloid membrane vesicle. (B) The fusion of two cup-shaped membrane vesicles. (C) The fusion of more membrane vesicles. (D) Vesicular budding analogous to endocytic invagination. Abbreviations: V, membrane vesicle; F, the place of fusion; R, ribosomes.
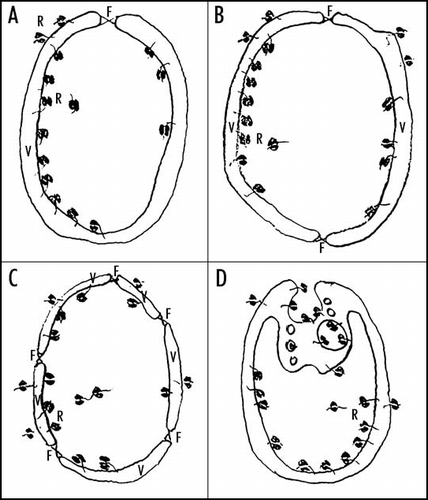
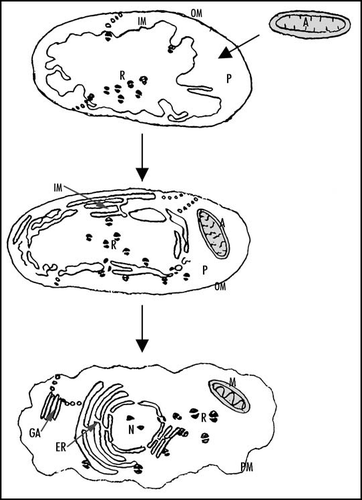
Figure 3 Hypothetical scenario for the origin of eukaryotic endomembranes. The pre-karyote is assumed to have been bounded by two membranes, inner (IM) and outer (OM). α-proteobacterial ancestors (A) of mitochondria (M) are proposed to have been parasites of pre-karyote periplasm (P). Endoplasmic reticulum (ER), nuclear membrane (N = nucleus) and Golgi apparatus (GA) are assumed to be derived from pre-karyote inner mebrane, while eukaryotic plasma membrane (PM) is assumed to be derived from pre-karyote outer membrane. It is suggested that ribosomes (R) were removed to pre-karyote periplasm (nowadays eukaryotic cytoplasm).