Figures & data
Figure 1.S. pombe telomeres are dissociated from the NE at M phase. (A) Telomere-binding proteins and their functions. (B) Telomeres are moderately clustered and are tethered to the NE in interphase. After entry into mitosis, telomeres are dissociated from the NE, and telomere clustering breaks down. For simplicity, only two chromosomes are shown.
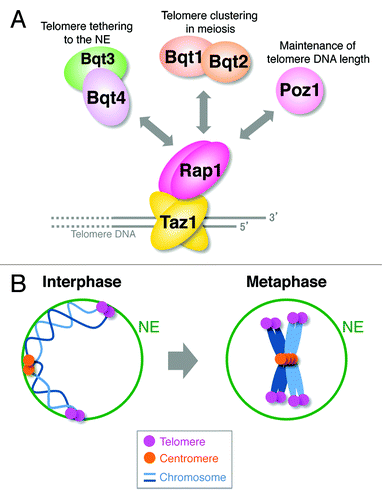
Figure 2. Rap1 is highly phosphorylated at M phase. (A) Band shifts of the Rap1 protein in M phase. Strain JK3332 (taz1–3HA nda3-KM311) was grown in YPD medium at 32°C to mid-log phase (0 h) and gradually arrested in early M phase by a temperature shift to 20°C (4–12 h). The whole-cell extracts at each time point were analyzed by immunoblotting with anti-Rap1 antibodies, anti-HA antibodies (for Taz1-HA), anti-Cdc13 (Cyclin-B) antibodies and anti-PSTAIR antibodies (for Cdc2). (B) Schematic illustration of the identified phosphorylation sites in Rap1. BRCT, BRACA1 C terminus; Myb, Myb-related HLH motif. C, constant during the cell cycle; E–M, early to mid M phase; L, late M phase. Thr378, Ser422 and Ser513 are phosphorylated by Cdc2.
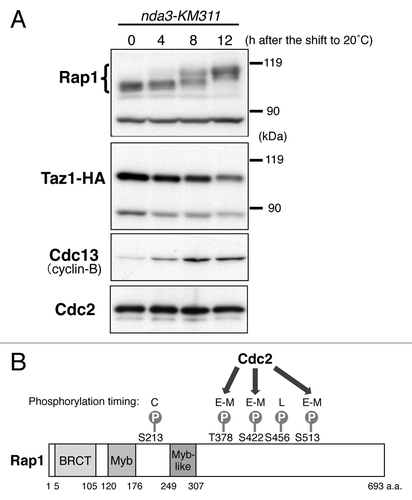
Figure 3. Model for regulation of telomere localization during the cell cycle in S. pombe. Telomeres are tethered to the NE via the Taz1-Rap1-Bqt4-Bqt3 association in interphase. During mitosis, Rap1 is phosphorylated. This phosphorylation, primarily by Cdc2, promotes the detachment of Rap1 from Bqt4 and thereby telomere dissociation from the NE. Transient telomere dissociation from the NE in mitosis is required for faithful chromosome segregation.
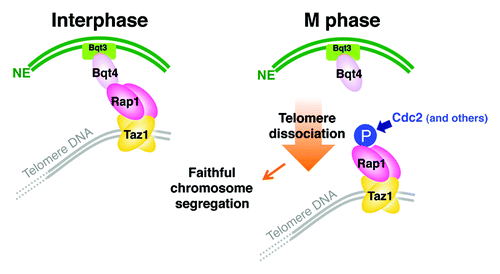
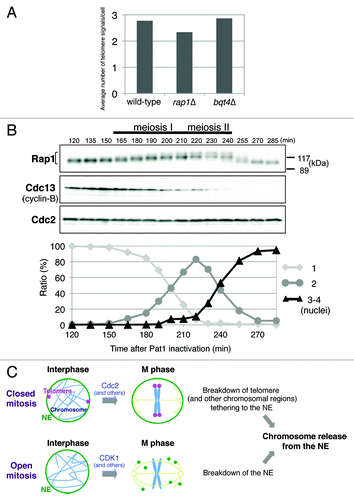
Figure 4. Conservation of chromosome release from the NE in eukaryotic cell division. (A) Mitotic telomere clustering in rap1∆ and bqt4∆ cells. Strains JK81 (wild-type, taz1-mCherry), JP836 (rap1∆) and JP842 (bqt4∆) were grown in YES medium at 30°C, and the numbers of Taz1-mCherry signals per cell were counted. More than 100 cells in G2 phase were analyzed for each strain. Note that S. pombe has three chromosomes and six telomeres. (B) Modification of the Rap1 protein in meiosis. Strain JP791 (h-/h-pat1–114/pat1–114 mat-Pc) was grown in YE medium to mid-log phase at 25°C and then transferred to EMM-N medium (supplemented with 1% glucose) at 25°C. After 6.5 h of incubation (G1 arrest), the temperature was shifted to 34°C to inactivate Pat1. Samples were taken every 10–15 min after 120 min of incubation at 34°C. The percentage of cells with 1, 2 or 3 –4 nuclei was determined by counting > 200 cells stained with DAPI (4',6-diamidino-2-phenylindole). The whole-cell extracts were analyzed by immunoblotting with anti-Rap1 antibodies, anti-Cdc13 (Cyclin-B) antibodies and anti-PSTAIR antibodies for Cdc2 (loading control). (C) Models for the regulation of closed and open mitoses. The breakdown of telomere tethering to the NE is triggered primarily by Cdc2 in closed mitosis in S. pombe. Dissociation of the other chromosomal regions from the NE may also be regulated by Cdc2. By contrast, breakdown of the NE triggered by CDK1 and other mitotic kinases occurs in open mitosis. Both induce the release of chromosomes from the NE, which likely leads to accurate chromosome segregation.