Figures & data
Figure 1 MicroRNA biogenesis. MiRNAs are mainly derived from RNA polymerase II transcribed genes. The primary transcript (pri-microRNA) is often polycistronic, containing several miRNas. In the nucleus, Drosha in conjunction with DRCG8 processes the pri-microRNAs into pre-microRNAs that are exported to the cytoplasm by Exportin5. In the cytoplasm, the mature miRNA is formed by the enzymatic activity of Dicer. The mature miRNAs is incorporated or “loaded” into the RISC (star shaped icon) that mediates miRNA-directed targeting of specific mRNAs.
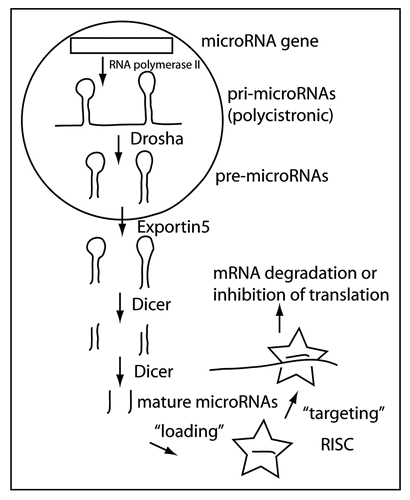
Figure 2 Disruption of regular scale pattern on tails of mice with epidermal specific loss of Dicer. Arrows indicate the regular lines separating scales on the tail of a control mouse (upper tail with hair). However, in mice with loss of Dicer function in the epidermis this regular pattern is destroyed (lower, darker and hairless tail).

Figure 3 Model for the role of microRNAs in the regulation of ‘tumor suppressor’ pathways in Drosophila. The miRNA bantam is a key target of the transcriptional regulator Yorkie. Yorkie is under tight control of the Hippo pathway that consists of merlin and expanded at the membrane integrating signals from an unknown transmembrane receptor. In certain cell types in Drosophila, expanded mRNA is regulated by miR-278. Warts and Mats transduce the signal from Merlin/Expanded and Hippo/Salvador. Warts and Mats are also elements of Fat/Dachsous signaling (another Drosophila tumor suppressor pathway) through Dachs. Fat also can interfere with canonical Wnt signaling and activate noncanonical Wnt signaling. This could theoretically occur via the inactivation of Yorkie and Bantam: one of Bantam's target mRNAs is Hid, which has been shown to inhibit canonical Wnt signaling.
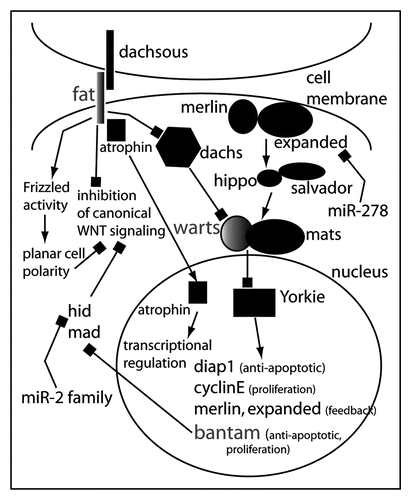
Table 1 Phenotypes of mice with a tissue-specific loss of Dicer
Table 2 The MicroRNA machinery and its association with human diseases