Figures & data
Figure 1 Proposed model for a role of B on cell-surface interactions driving symbiosome development. Golgi derived vesicles (V) loaded with peribacteroid components, including lectin-like glycoprotein PsNLEC 1 (vc), are targeted to the symbiosome compartment. In the presence of borate anion (represented as B−) (+B panel), RGII-glycoproteins (GP) are stabilized on the inner face of vesicle membrane (VM). Then, it is able to interact with PsNLEC 1, which also anchors to the LPS of the bacteroid (B) reinforcing vesicle fusion. The triple association PBM-PsNLEC 1-bacteroid allows juvenile bacteroid differentiation into a mature N2-fixing form. In the absence of borate (−B panel), the vesicle membrane is not decorated with RGII-antigens; therefore it is unable to interact with PsNLEC 1. Consequently vesicle fusion becomes unstable and subsequent bacteroid development is arrested at a juvenile immature non N2-fixing stage.
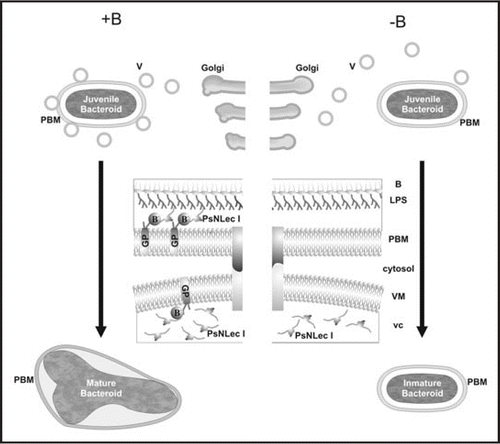
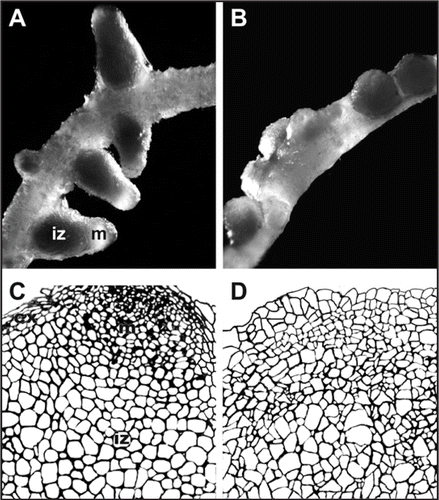
Figure 2 Effects of B on nodule organogenesis. (A) Pea root nodulated with sufficient B showing nodules with colourless apical nodule meristem (m) and dark central infected zones (iz). (B) Pea root nodulated in the absence of B, showing nodules with a tumour-like appearance when different development zones are undistinguishable. (C) Schematic illustration of B-sufficient nodule tissue organization indicating the apical meristem (m), the infected zone (iz), and the nodule cortex (cx). (D) Schematic illustration of highly disorganized B-deficient nodule structure with not clear development of different nodule tissues. (C and D) are based on (previously published studies, refs. Citation20 and Citation21).