
Abstract
Apoptosis or programmed cell death applies as a protected way to remove unwanted cells and plays a determinative role in many important biological processes such as normal development and tissue homeostasis in multicellular organisms. The main purpose of studies in this field is to focus on understanding the related regulatory mechanisms and novel approaches to the discovery of anticancer drugs. Among the apoptotic messenger molecules, the key roles of sphingosine and ceramide as the backbone of sphingolipids were considered in many pathways of apoptosis. It has been shown that ceramide plays an important regulatory role in apoptosis and its increased cytoplasmic levels lead to programmed cell death. Ceramide biosynthesis and biodegradation occur via several different enzymes they can be affected by many external factors. Instead, phosphorylated form sphingosine is an important biomarker for cell proliferation and differentiation. Phosphatases, kinases, and proteases are the main operators for ceramide messaging and they can affect different pathways of apoptosis directly or indirectly. Sphingosine is an extracellular ligand for G-protein receptors and an intracellular second messenger to promote cell survival. Many different types of current chemotherapy drugs and some understudy compounds for cancer can at least affect one of the ceramide metabolizing enzymes. Serine palmitoyltransferases, sphingomyelinase, ceramidase, and glucosylceramide synthase are the most important enzymes involved in sphingolipid metabolism have been the discussed in many studies. Consequently, sphingolipids and their related metabolizing enzymes introduced as new pharmacological targets will be helpful in anticancer drug development.
Abbreviations:
- AIP
- apoptosis-inducing factor
- BAD
- Bcl-2 associated death promoter
- BAK
- Bcl-2-antagonist/killer
- BAX
- bcl-2 associated X Protein
- BID
- BH3 interacting domain death
- COX
- cyclooxygenase
- D-MAPP
- dimethylallyl pyrophosphate
- ERK
- extracellular signal-regulated kinase
- FADD
- Fas-associated protein with death domain
- FAN
- factor associated neutral sphingomyelinase
- GPCR
- G protein-coupled receptor
- IAP
- Inhibitor of apoptosis protein
- IL-1
- Interleukin-1
- MAPP
- myristoylamino-phenylpropanol
- mTOR
- mechanistic target of rapamycin
- NF-қB
- nuclear factor-kappa-B
- PDGF
- Platelet-derived growth factor
- PI3K
- phosphoinositide 3-kinase
- PLC
- phospholipase C
- SMAC
- second mitochondria-derived activator of caspases
- SPT
- serine palmitoyltransferase
- TLR
- toll-like receptor
- TNF
- tumor necrosis factor
- TNFR
- TNF receptor
- TRADD
- TNF receptor type 1-associated death domain
- TRAIL
- TNF related apoptosis-inducing ligand
- TRAMP
- TNF receptor-related apoptosis-mediating protein
- UDP
- uridine diphosphate
1 Introduction
In the past few years, lipids were mainly considered for their role in maintaining the structure of the membrane and protecting it from environmental stresses in cellular-molecular studies. The findings of the last two decades have greatly changed this concept, and today many lipids play a key role as secondary messengers in regulating different cellular processes, including inflammation, fever, survival, cell proliferation, cell division, differentiation, aging and cell death [Citation1]. Sphingolipids have been considered as the most important profile of the lipid messengers, and the factors affecting them have been the subject of various studies [Citation2]. They are a group of 18-carbon alcohol-based lipids, namely, sphingosine, and are made in an endoplasmic system [Citation1].
The changes that sphingosine undergoes in the influence of various enzymatic systems will ultimately lead to the formation of sphingolipids with an important structural role in the cytoplasmic membrane as well as bioactive mediators in the regulation of cell homeostasis [Citation2]. Contrary to the construction variability of the sphingolipids, functional enzymes in the fabrication, transformation, and destruction of these compounds are common systems in cellular metabolism [Citation3].
The occurrence of apoptosis or programmed cell death as one of the most important processes for the preservation of homeostasis in multicellular organisms is commonly used to remove unwanted or unnecessary cells and interferes with many mechanisms of the immune system or diseases [Citation4]. Apoptosis plays a vital role in important biological processes, such as natural development, homeostasis, removal of damaged or infected cells, or activated immune cells against self-administered antigens. The incidence of many autoimmune diseases, cancers, and viral infections are results of poor performance or inhibition of apoptosis [Citation5]. The lack of equilibrium in the incidence of this phenomenon, both decreasingly and incrementally, causes cancer or neurodegenerative disorder such as Parkinson's and Alzheimer’s disease [Citation4,Citation5] .
The importance of protecting cell processes from apoptosis as well as its inducing factors and, finally, the balance between these factors are so considerable. The evaluation of stimulus or inhibitor factors and messenger pathways in the occurrence of apoptosis regarded as one of the most attractive and applicable categories in various fields of biology [Citation2,Citation3] .
Among the factors regulating this physiological (and sometimes pathologic) process, some of the sphingolipid molecules have played a determining role [Citation2]. It has been shown that the phosphorylated sphingosine is one of the most potent inhibitors of apoptosis and mitogen stimuli, and the phosphorylated form of ceramide as a sphingosine precursor is a great inducer of apoptosis [Citation1,Citation6] . In addition, it has been demonstrated that the balance between the sphingosine and ceramide, as a determining factor in the fate of the cell, utilizes various operators in sending a message of life or death to the cell [Citation3].
In the recent years, the factors enhancing or inhibiting ceramide operators and sphingolipids metabolizing enzymes, regarded as novel pharmacological targets for controlling apoptosis [Citation6]. Other cellular processes dependent on these messengers have been the subject of discussion of various biological studies [Citation7].
Considering the extent and complexity of ceramide and sphingosine operators and metabolizing systems as one of the most important apoptosis regulators and the introduction of some drugs affecting these pathways into clinical trials, further study in this area will be hope in treating many related disorders, including cancer [Citation3,Citation8] .
2 The cellular apoptosis pathway
The inhibitors of apoptosis proteins (IAPs) and apoptosis-inducing proteins (AIPs) play important roles in the regulation of planned cell death. The occurrence or non-occurrence of a apoptosis, like any other physiological phenomenon, results from the balancing of a large number of messenger molecules with the effects on IAPs and AIPs [Citation5,Citation9] . These messenger pathways, by activating a large family of proteases called initiator caspases, have resulted in the activation of functional caspases, and ultimately functional caspases also by compacting and fragmenting the chromatin, cause the planed cell death [Citation4].
Among the proteolytic enzymes, caspases are a group of proteins called cysteine proteases. The induction of initiating caspases, results in the breakdown of inactive dimer pre-caspases and their conversion into active and functional caspases [Citation5].
Although two intrinsic (mitochondria-dependent) and extrinsic (dependent on cell death membrane receptors) have been identified as the main pathways for induction of apoptosis, other mechanisms, such as the endoplasmic and lysosomal pathway, have been recently reported [Citation4,Citation10] . The p53 molecule, as one of the most important tumor suppressors, plays a central role in stimulating the internal pathways of apoptosis and the damage to DNA is the critical factor in its action [Citation11].
In the intrinsic pathway, factors such as environmental stress, heat, hypoxia, lack of growth factors, infections from intracellular masses and caspase-8 which activated in the external pathway of apoptosis results in increased expression of pro-apoptotic proteins of the B-cell lymphoma family such as BAX, BAK, BID, BAD and nitric oxide and attenuation of the anti-apoptosis proteins such as Bcl-2 and Bcl-XL [Citation5,Citation9,Citation12] . The total of these events leads to increased permeability of the mitochondrial membrane and the formation of pores in its surface, resulting in the release of factors such as Cytochrome C and SMAC from mitochondria into the cytoplasm [Citation13]. The release of these agents by generating the apoptosome complex activates Caspase-9, and the activation of Caspase-9 induces caspase 3 which leads to apoptosis [Citation14].
In the extrinsic pathway, the activation of receptors from the TNF family such as TNFR-1, CD95 (Fas), TRAMP, TRAIL-1 and 2 in the cell membrane resulted in the binding of these receptors from its intracellular sequences to ligands such as TNF-α, lymphotoxin, and Fas ligand finally, intermediaries like TRADD and FADD produce, which are Caspase-8 activators [Citation4]. Caspase-8 also causes apoptosis with increased expression of pro-apoptotic proteins of the B-cell family and activation of functional caspases [Citation9]. In the lysosomal pathway of apoptosis, the release of lysosomal proteases including cathepsin B, acid sphingomyelinase, acid ceramidase, etc., apart from being able to lead the cell to autophagy, will also stimulate the onset of apoptosis [Citation10,Citation15] .
The endoplasmic reticulum is one of the major apoptosis regulating cellular organelles that plays the key role in this important biological event by making many apoptotic or antiapoptotic stimulants, including Bcl family proteins, death receptor ligands, and the release of calcium into the cytoplasm [Citation2,Citation5] . In addition, the newly synthesized (De Novo) ceramide, as one of the most important messenger phospholipids in cellular death, occurs in the endoplasmic reticulum. Golgi apparatus also plays a critical role in protecting cells from the apoptotic process by making glucosylceramides and P-glycoproteins [Citation1].
3 Structure and role of sphingolipids in cellular physiology
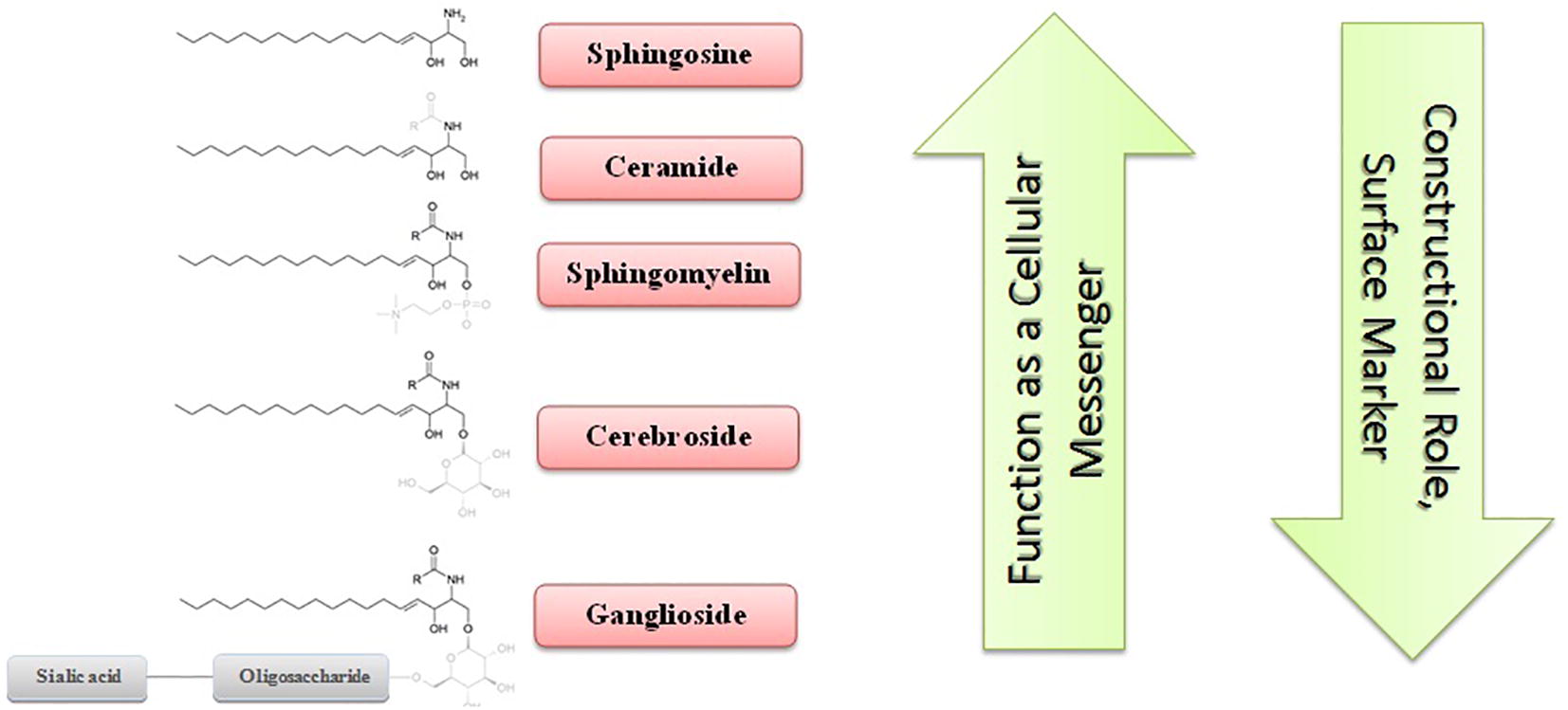
Sphingolipids are a class of lipids with sphingosine as a complex amino alcohol core containing 18 carbons, which together with glycerolipids and sterols, form the major part of the cell membrane structure. Ceramide as ubiquitous sphingolipids is composed of a sphingosine, which is amide-bonded to a fatty acyl chain with different numbers of carbons, ranging from 14 to 26 [Citation16]. Ceramides are one of the major components of the cell membrane and play a key role in protecting the cell from environmental stress [Citation11]. The addition of phosphoethanolamine, monosaccharide or oligosaccharide together with sialic acid to ceramide leads to the formation of sphingomyelin, cerebrosides, and ganglioside, which, apart from the structural importance, they play a role as biological markers and the binding agent of extracellular ligands to the receptor [Citation3,Citation17] .
However, over the past two decades, it has been shown that sphingolipids, especially ceramides and sphingosine, in their phosphorylated forms as messenger molecules, are crucial mediators of many physiological and pathological cellular processes, and the balance of their levels determines the fate of the cell [Citation1,Citation3] . Configuring processes of proliferation, cell division, fever and inflammation, aging, lipid, and sugar metabolism regulation, and many other things are also affected by these messenger molecules. In contrast, more complex sphingolipids have a constructional role and, except for a few cases, do not function as a secondary messenger [Citation18].
shows the molecular structure of important sphingolipids and compares their structural and messenger roles.
Fig. 1 The structure of the important sphingolipids and comparing their messenger and structural roles. Simple sphingolipids have been considered mostly as messengers and complex molecules have been principally notable for their structural role.
4 Sphingolipids as messenger molecules in cell survival
In recent years the results of numerous studies have shown the determinant role of ceramides and total cell sphingosine (in the phosphorylated form) in the formation of a death or cell survival message [Citation3]. It has been shown that if this balance changes in any way towards increasing total cellular ceramide, the various pathways of the apoptosis occurrence, including the internal, external and common pathway, will be activated quickly and the cell will soon be directed to the planned death [Citation2,Citation18] . Conversely, the cleavage of ceramides in the cytoplasm by increasing the activity of various isoforms of ceramidase enzymes leads to an increase in the levels of intracellular sphingosine [Citation19]. Sphingosine, by mediating environmental growth factors and stimulant cytokines of proliferation and division, is subjected to the action of the sphingosine-1-kinase. The phosphorylated sphingosine directly and indirectly (through the GPCR receptor) stimulates the factors involved in cell division, such as PI3K and PLC [Citation6,Citation20] .
shows a summary of the messaging paths of ceramide and sphingosine in determining the fate of the cell.
Fig. 2 The balance between total cellular phosphorylated sphingosine and ceramide levels as a regulator factor of cell survival or death message. Activating the main pathways of ceramide production, by increasing the total cellular ceramide levels, trigger the release of cytochrome C (Cyt C), which act on downstream activation of FADD (Factor associated cell death) and most importantly, increase the activity and expression of ceramide operators include CAPKs (Ceramide-activated protein kinases), cathepsin B and PP1 (protein phosphatase 1) and eventually activate caspases and induce apoptosis. Increasing the destruction of ceramide and its transformation into sphingosine and the activity of the sphingosine kinase also result in an enhancement in the intracellular content of the phosphorylated sphingosine and this mediator, by its G-protein coupled receptor receptors and by increase of the expression of PI3K (phosphoinositide 3-kinase), PLC (Phospholipase C), ERK (extracellular signal-regulated kinase), and COX II (cyclooxygenase II) propagate the message of life and division.
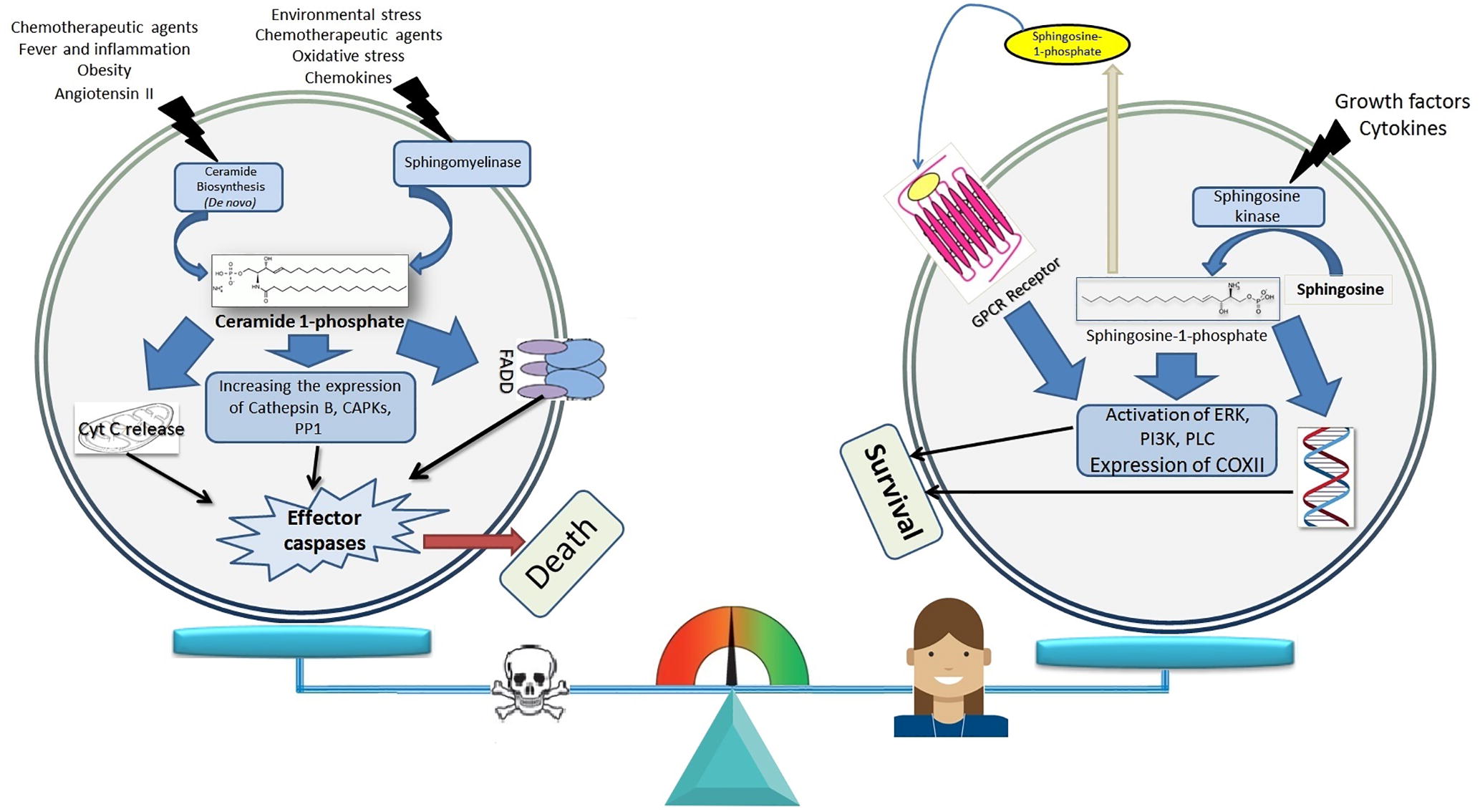
It has been shown that TNF induces apoptosis in tumor cells by increasing the production of ceramide, and in this way, ceramide does not require p53 [Citation11]. The addition of exogenous ceramide also induces programmed cell death and ceramide generation is a key controller of apoptosis [Citation7,Citation21] .
5 The main pathways of the sphingolipid metabolism as a new target for chemotherapy
The ceramide signaling pathway is known as a principal apoptosis-inducing factor in diverse tumor cells [Citation6]. In recent decades, the elements contributing to the exogenous ceramide levels, their operators and the factors influencing its metabolism have been the subject of numerous studies [Citation7]. Hence, consideration of enzyme systems which involved in the ceramide and sphingosine metabolism as main sphingolipids in the cell survival or cell death, and the mechanism of action of their effectors will be very helpful in this regard [Citation17].
Many of the ceramide catabolism inhibitors, as well as its anabolism boosters, are passing clinical phases as tumor chemotherapy drugs [Citation22]. In addition, the production of ceramide and the potentiation of its operators have been proposed as a secondary mechanism of anti-tumor activity of many chemotherapy agents and radiotherapy [Citation19].
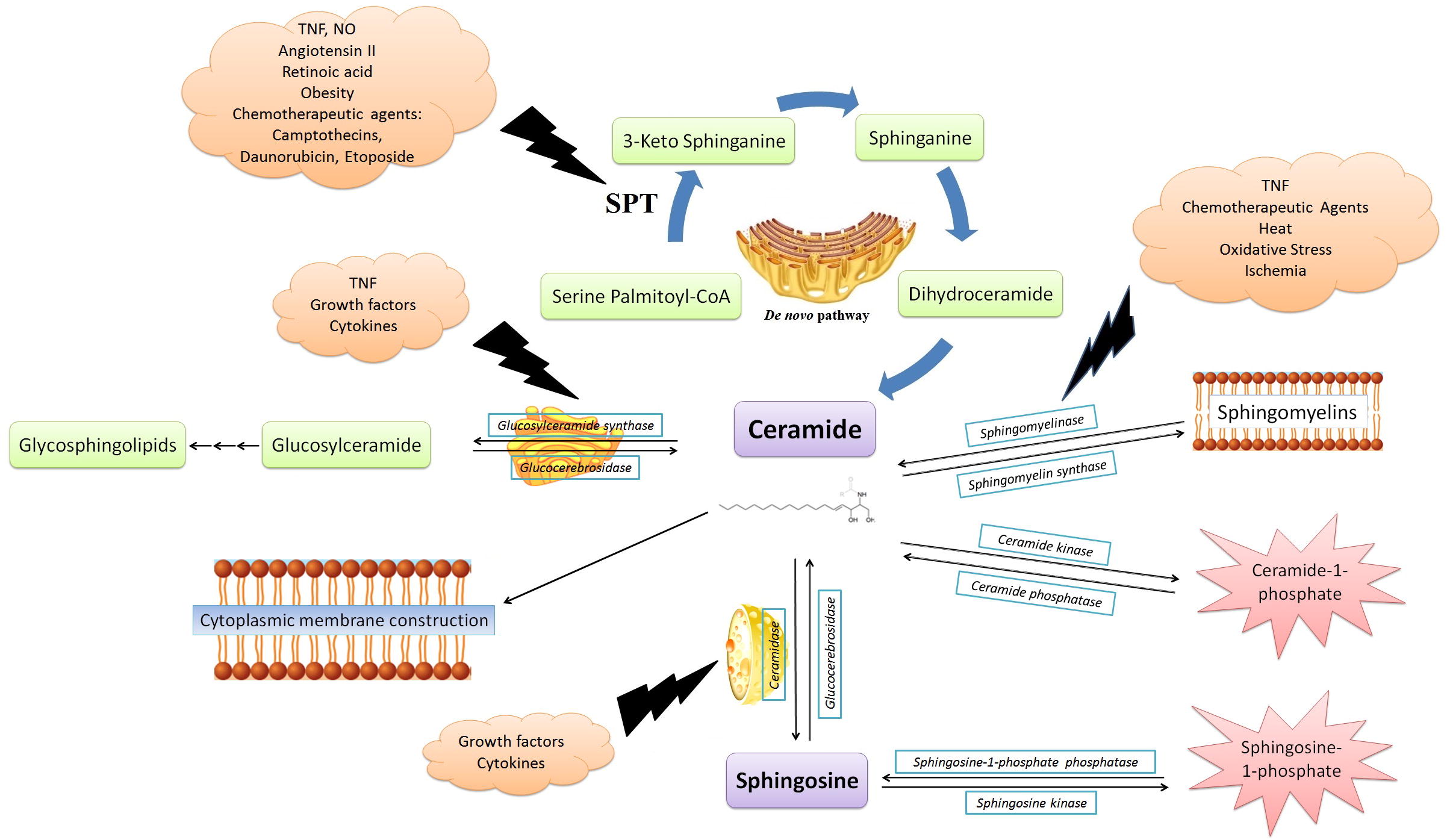
In , some examples of pharmacological agents which affect important enzyme systems involved in the sphingolipid metabolism are presented. also shows a summary of different pathways of intracellular ceramide metabolism.
Table 1 The factors affecting important enzymes involved in ceramide metabolism.
Fig. 3 The main routes of ceramide metabolism. The most important pathway for the ceramide biosynthesis is the De novo pathway, in which the first step of the coupling of palmitic acid to the coenzyme A with the D-serine amino acid occurs by the SPT enzyme. After that, in the next three steps, the ceramide will eventually be obtained. Ceramide can be converted to sphingomyelin, or diverse isoforms of sphingomyelinase break sphingomyelin in the cytoplasmic membrane into ceramide. The glucosylceramide synthase enzyme in the Golgi apparatus can glucosylate the ceramide present within the cytoplasm and finally produce glycosphingolipids in several steps. Glucocerebrosidase is another enzyme that can reverse the ceramide, unlike the previous one. The breakdown of ceramide by various isoforms of ceramidase and its transformation into sphingosine and free fatty acid creates another event that can affect ceramide. In contrast, Ceramide synthase produces ceramide by connecting the sphingosine to free fatty acid. The phosphorylation of ceramides and sphingosine in the cytoplasm has led them to become an active form of their biological mediators, and this incident makes them a secondary messengers. Finally, free ceramide in the cytoplasm can be used to make cytoplasmic membranes.
6 De novo pathway
The onset of the biosynthesis of cytoplasmic ceramide and a major part of other sphingolipids, also known as the De novo Ceramide synthesis, is from the endoplasmic reticulum and results in the formation of ceramides from palmitoyl-CoA and serine in five steps [Citation17]. The first step and the most important stage of ceramide biosynthesis, which is the limiting reaction speed, is the binding of serine and palmitic acid to coenzyme A, catalyzed by serine palmitoyltransferase (SPT) and results in 3-ketosphinganine generation [Citation1]. After that, 3-ketosphinganine undergoes resuscitation, N-acetylation, and desaturation and eventually, after the addition of acyl-coenzyme A with a different number of carbons, ceramide is obtained with various numbers of carbons of the acyl group (6–24) [Citation17].
It has been shown that inhibition of ceramide construction by suppression of this pathway with specific SPT inhibitors such as Fumonisin II, has reduced the response to tumor chemotherapy and, conversely, the mutation in this enzyme and its high basal activity, will result in an inherent discomfort of the sensory nerves [Citation23]. Different biological agents such as nitric oxide, TNF, angiotensin II, the frequency of palmitic acid reserves, some conditions such as obesity, as well as the presence of pharmacological agents such as retinoic acid derivatives, camptothecin, daunorubicin have been introduced as the stimulus of SPT activity [Citation1,Citation17] .
7 Sphingomyelinase pathway
The ceramide which made in the endoplasmic reticulum can be introduced into the Golgi apparatus by its own carriers and subject to different isoforms of the sphingomyelin synthase enzyme and convert to sphingomyelin with a similar number of carbons to that of the original ceramide. This happens by converting phosphorylcholine to diacylglycerol [Citation31]. The product from this reaction is mainly used for the manufacture of myelin of the cell membrane of neurons or in the presence of the sphingomyelinase enzyme convert to ceramide [Citation32].
The acidic form of sphingomyelinase is one of the most important isoforms of this lysosomal enzyme that performs the degradation and conversion of cytoplasmic membrane sphingomyelin to ceramide [Citation26]. Therefore, its strengthening has become a target for anti-tumor drugs. The neutral and alkaline isoforms of sphingomyelinase are mainly present in the cytoplasm and are often magnesium and zinc-dependent, and the abundance of these heavy metals is also a factor in the enhancement of these enzymatic systems [Citation33]. Suppression of this pathway in cancer cells has been introduced as a resistance mechanism to some chemotherapy agents [Citation26].
Variety of sphingomyelin to ceramide converting enzymes leads to different biological responses from influential factors. Specific biologic factors such as interferon gamma, interleukin-1, heat, oxidative stress, ischemia/reperfusion stress, and factors associated with neutral sphingomyelinase (FAN) and pharmacological agents such as vitamin D3, doxorubicin, mitoxantrone and silibinin been proposed as an enhancer for varied numbers of sphingomyelinase isoforms [Citation24,Citation25,Citation30] . These drugs either have anti-tumor effects, and increased ceramide production through induction of sphingomyelinase is a secondary mechanism for their effects or by enhancing the production of ceramide from this route, the probability of potential anti-tumor or tumor-preventative effects for them is posed.
On the other hand, in some studies, sphingomyelinase inhibitors also have protective effects on apoptosis in different cells. It has been demonstrated that antidepressants such as desipramine and amitriptyline, as inhibitors of acid sphingomyelinase, inhibit apoptosis, have a protective role in the heart cells and protect liver cells from fibrosis [Citation26].
8 Ceramidase pathway
The ceramide present within the cytoplasm or in the lysosome can be broken down under the influence of various isoforms of the ceramidase enzyme and converted to the sphingosine and its free fatty acid [Citation16]. So that not only the cell will be saved from the death mediated by ceramide, but its sphingosine base also converts to phosphorylated form, and the cells are directed to the survival and mitogenic condition [Citation34]. Consequently, inhibitors of this enzyme can be considered as tumor suppressor agents [Citation35].
Among the three types of ceramidase (acid, alkaline, and neutralized), the acidic form which presents in the lysosome has been studied as the main form in several investigations [Citation19]. Endogenous and pharmacologic agents such as TNF, IL-1, Platelet-derived growth factor (PDGF) and etoposide, tamoxifen and dacarbazine are known as ceramidase inhibitors and induce apoptosis with ceramide accumulation from this pathway. The mutation in this enzyme system and its lack of response to dacarbazine, have been introduced as a factor in the resistance of tumor cells to this drug [Citation35]. We have previously shown that hesperidin derivatives by inhibiting this enzyme are able to induce the ceramide-dependent apoptosis in HT-144 melanoma cancer cells [Citation36].
The importance of ceramides in the cancer biology is in the way that its selective inhibitors are developing and some of them, such as MAPP and B-13, enter the clinical phases of the treatment of some tumors, such as a malignant neoplasm of the prostate [Citation22,Citation37] . Regarding the presence of the most important isoform of the ceramidase, namely acid ceramidase in the lysosome, access and change the structure-activity relationship of drugs in order to increase their efficacy have been challenging issues in related studies on this field [Citation8,Citation28] .
9 Glucosylceramide synthase pathway
The entrance of ceramide to the Golgi apparatus is considered as the gateway for the manufacture of glycosphingolipids [Citation29]. In this case, the first step is the addition of glucose from UDP-glucose to ceramide by glucosylceramide synthase enzyme and glucosylated ceramide production [Citation17]. In the next steps, various products of glycosphingolipids, which are mainly structural and cellular markers, are derived from this mediator or additional glucosylated ceramides in the inverse pathway of the reaction are converted by cerebrosidase into its constructive ceramide [Citation3]. Thus, the activity of glucosylceramide synthase is a cell rescuer from ceramide-induced apoptosis and a way to chemotherapy resistance [Citation38].
Growth factors are mainly amplifying this pathway due to the cell's need for more bulking glycosphingolipids in the cell membrane construction [Citation39]. However, the role of the inhibitors of this pathway to ceramide elimination has been low in cell death studies, and glucosylceramide synthase inhibitors such as GZ667161 have been used in some neurological disorders [Citation29]. The previous studies have been confirmed that some of the bioflavonoid derivatives such as silibinin and hesperidin by inhibiting this enzyme, allow the accumulation of ceramide in tumor cells [Citation30,Citation36,Citation40] .
10 Ceramide signaling pathways
Phosphorylated ceramides use multiple operators to announce the programmed cell death orders. In fact, the ceramide messaging route is located in the upstream of many other messenger molecules and is significant in this regard [Citation21]. Of course, it should be noted that all three main pathways for apoptosis are directly induced by the ceramide pathway, although ceramide operators seem to play a more important role in terms of multiplicity and efficiency [Citation41,Citation42] . In some cases, ceramide has been shown to be capable of inducing programmed cell death without activating caspases.
Generally, the ceramide messenger path operators are divided into three categories of kinases, phosphatase, and proteases, which we will discuss each of them in the following [Citation41].
Ceramide-activated protein phosphatases (CAPPs) are the most important mediator for ceramide signaling pathways, with two major elements: proteins phosphatase 2 A (PP2A) and protein phosphatase (PP1). PP2A deactivates the main cell survival and division mediators such as Akt, c-JUN, PKCα, and BCL2, thus shutting down the propagation engine and cellular life [Citation1,Citation42] .
Akt or protein kinase B is located in the intersection of cell metabolism and is directly in a relationship with mTOR, which controls the metabolism and vitality of many of the cells throughout the body, especially the immune-system cells. Some studies point to the fact that the activation of caspase-9 and BAD is possible only with the Akt dephosphorylation [Citation2].
The c-JUN phosphorylated form, together with cFos, is responsible for the construction of many cell cycle transcription factors and it has been shown that this mediator in association with NF-κB is able to save the cell from a planned death induced by TNF or UV waves [Citation43]. It has already been revealed that ceramide prepares the cell for apoptosis by turning off c-JUN.
Among the various PKC subtypes, at least two of them are influenced by the ceramide messaging pathway: PKCα and PKCζ. PKCα is one of the common types of PKC family that requires calcium liberation in cytoplasm and diacylglycerol in a cytoplasmic membrane for signaling, in order to activate ultimately many cell survival factors, including p38 and MAP-kinase [Citation42]. Ceramide by dephosphorylating this sub-group suppresses all related pathways. On the other hand, ceramide, by increasing the activity of PKCζ, delays the rearrangement of the Bcl-x gene family as one of the most critical apoptotic suppressors, thus causing cellular death [Citation2].
PP1, The second group of downstream phosphatases which are dependent on the ceramide pathway, play an important role in controlling of the two families of cell cycle regulator, SR and Rb proteins [Citation3]. SR proteins (proteins with repetitive sequences of arginine and serine) in their non-phosphorylated form facilitating the rearrangement of caspase-9 and Bcl-x inhibition, leading to apoptosis [Citation42]. Retinoblastoma protein (Rb) is another tumor suppressor that only in a phosphorylated moiety allows to passing cell through a cell cycle phase to another, and, naturally, its dephosphorylation will stop the cell cycle [Citation1].
The kinases are the second group of the factors influenced by ceramide and interestingly; the downstream phosphorylation of this pathway is mainly due to the suppression of the messenger’s cascade [Citation42]. Ceramide-activated protein kinases (CAPKs) are the most considerable kinases activated by ceramide and inhibit GTPase activity by phosphorylation of the Rat sarcoma protein (RAS) [Citation3]. Subsequently, extracellular signal-regulated kinases (ERK) pathways are inhibited as one of the main pathways for DNA replication. On the other hand, the rapidly accelerated fibrosarcoma (Raf) Factor is also activated by another group of CAPKs, which carries the message of ceramide in inflammation induced by Interleukin-1 [Citation1].
Proteases are the third main group of ceramide-1-phosphate operators, among which cathepsin B is the most prominent member of this group. After activation of cathepsin B by ceramide, BID is broken, and truncated BID (tBID) by releasing cytochrome C from the perforations created in the mitochondria will act as an activator of caspases in apoptosis [Citation2,Citation14] .
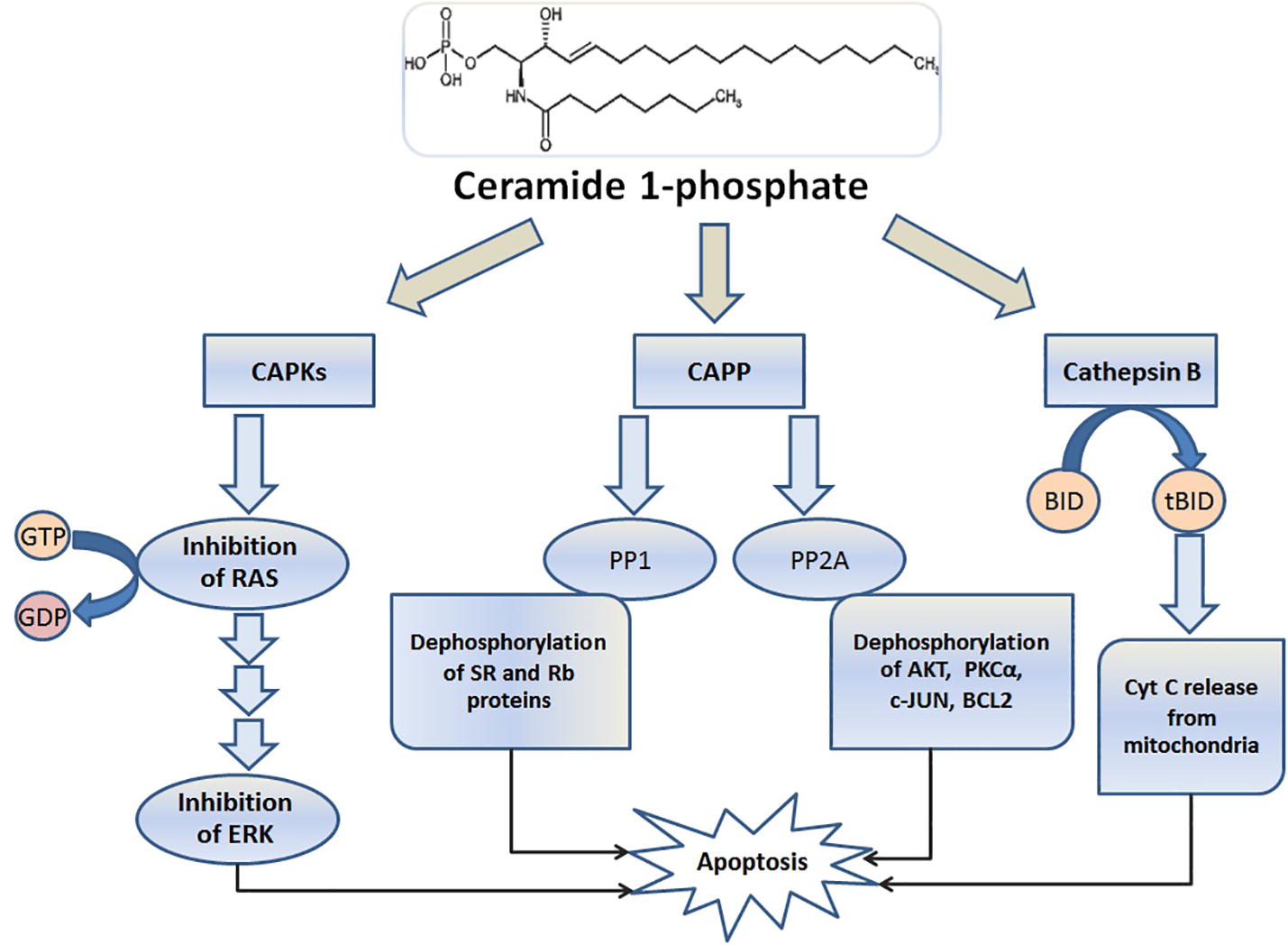
shows the summary of cellular pathways affected by ceramide operators in apoptosis.
Fig. 4 The major effectors of ceramide. The phosphorylated ceramide inside the cytoplasm as a secondary messenger serves of the three main categories of operators for sending death message to the cell. In the kinase group, Ceramide-activated protein kinases (CAPKs) are the most important kinases that activated by ceramide and CAPKs also inhibit Rat sarcoma (RAS) phosphorylation of its GTPase activity. In this way, the pathway of extracellular signal-regulated kinases (ERK) is inhibited as one of the main pathways for proliferation and cell division. Cathepsin B (as a protease) and ceramide produce a truncated BID (tBID), and the tBID activates the caspases dependent apoptosis by releasing cytochrome C from the pore in the mitochondria. Ceramide-activated protein phosphatases (CAPPs) is the third group of ceramide operators that their two main groups, Protein phosphatase 2A (PP2A) and Protein phosphatase 1 (PP1), dephosphorylate many of the factors, which involved in maintaining the cell survival.
11 The cell signaling pathway of sphingosine
The sphingosine derived from the decomposition of ceramide by stimulating external factors such as growth promoters and some cytokines, as well as the downstream activation of the TNF receptor, is affected by the phosphodiesterase 1-kinase and acts as a bioactive mediator [Citation18]. Phosphorylated sphingosine, by affecting some of the regulatory factors in the cytoplasm and with its intrinsic effect on related G protein-coupled receptors (GPCRs) as a cell membrane surface receptors, (mainly in opposition to the pathway of ceramide) will direct the cell to proliferation and differentiation [Citation6]. Hence, more expression of sphingosine 1-kinase in high-proliferation cells such as immune and mucosal cells is expected [Citation1].
The key role of the sphingosine and its receptors have been identified in many biological phenomena such as angiogenesis, vascular permeability, and chemotaxis of B and T lymphocytes, as well as TLR receptor-dependent immunosuppression, have been identified [Citation6]. Moreover, the reinforcement of this messaging pathway has been introduced as a mechanism for resistance to some chemotherapy agents in cancer cells.
Phosphorylated sphingosine by directly activating COX II, ERK, and NF-κB, and deactivating BAX and Caspase-3, leads to inhibition of apoptosis and induction of cell division [Citation15]. This mediator can also withdraw from the cell and act as an autocrine or paracrine on its GPCR and depending on the type of receptor, activates various messaging paths. They include G12/G13, Gq/G1, Gs and Gi/Go and the multiplicity and diversity of these receptors justify some of the contradictory effects observed from the sphingosine in cellular life [Citation6].
12 The role of other sphingolipids in apoptosis
As noted above, glycosphingolipids with a complex sugar structure mainly play a composition role for the cell membrane, and apart from this, they act as cellular markers and receptors. For example, the toxin of some microbial agents uses gangliosides as a pathogen binding agent to enter the cell [Citation44].
In accordance with the subject, it has been seen that in some regions of the brain during development, Ganglioside GM3 and GD3 were significantly more pronounced than other regions, which indicates their potential role in the development of the nervous system [Citation45,Citation46] .
While monoclonal antibodies against ganglioside GD2 had beneficial effects in the treatment of melanoma, it has been shown that GD3 as one of the downstream products of glycosylation of ceramide induces apoptosis and plays an essential role in the removal of unwanted cells during evolution [Citation39,Citation47] . whereas high levels of GM3 associated with Parkinson's disease with unknown origin, some studies have exhibited that this factor, by following neurotrophic agents, has a protective and increasing role for neuronal plasticity, and its derivatives are known as effective agents for the treatment of Alzheimer’s and Huntington’s Disease [Citation48Citation[49]–Citation50] . In addition to the above, in many studies, glycosphingolipids played a regulatory and pivotal role as a secondary messenger in the process of insulin resistance and fatty liver induction [Citation51Citation[52]–Citation53] .
13 Conclusion
The occurrence of apoptosis is one of the main protective methods for porcelain organisms in eliminating excess cells and/or avoiding cell cycle regulatory processes and preventing them from entering the neoplasm phase. The subcellular balance of the sphingosine/ceramide cytoplasm as the most significant apoptotic regulator determines the fate of the cell, and drug interventions on the enhancing or facilitating factors of each of them will control the transform of the cell to death or life. Increasing the knowledge of molecular biology in the context of metabolic enzyme systems, Intracellular downstream operators, as well as their external influential factors, will provide a new insight in the treatment of diseases resulting from apoptosis disorders, such as cancer, diabetes, Parkinson's and Alzheimer's disease.
References
- B.J.PettusC.E.ChalfantY.A.HannunCeramide in apoptosis: an overview and current perspectivesBiochim Biophys Acta15852002114125
- B.OgretmenY.A.HannunBiologically active sphingolipids in cancer pathogenesis and treatmentNat Rev Cancer42004604616
- C.P.ReynoldsaB.J.MaureraR.N.KolesnickCeramide synthesis and metabolism as a target for cancer therapyCancer Lett2062004169180
- L.OuyangZ.ShiS.ZhaoF.T.WangT.T.Zhouet al.Programmed cell death pathways in cancer: a review of apoptosis, autophagy and programmed necrosisCell Prolif452012487498
- S.ElmoreApoptosis: a review of programmed cell deathToxicol Pathol352007495516
- N.J.PyneS.PyneSphingosine 1-phosphate, and cancerNat Rev Cancer102010489503
- T.H.BeckhamJ.C.ChengS.T.MarrisonJ.S.NorrisX.LiuInterdiction of sphingolipid metabolism to improve standard cancer therapiesAdv Cancer Res1172013136
- D.PizziraniC.PaqliucaN.RealiniD.BranduardiG.BottegoniDiscovery of a new class of highly potent inhibitors of acid ceramidase: synthesis and structure-activity relationship (SAR)J Med Chem56201335183530
- X.M.YinSignal transduction mediated by Bid, a pro-death Bcl-2 family proteins, connects the death receptor and mitochondria apoptosis pathwaysCell Res102000161167
- M.E.GuicciardiM.LeistG.J.GoresLysosomes in cell deathOncogene23200428812890
- R.Hage-SleimanM.O.EsmerianH.KobeissyG.Dbaibop53 and ceramide as collaborators in the stress responseInt J Mol Sci14201349825012
- G.DewsonR.M.KluckMechanisms by which Bak and Bax permeabilise mitochondria during apoptosisJ Cell Sci122200928012808
- J.E.ChipukD.R.GreenHow do BCL-2 proteins induce mitochondrial outer membrane permeabilization?Trends Cell Biol182008157164
- S.J.KorsmeyerM.C.WeiM.SaitoS.WeilerK.J.OhP.H.SchlesingerPro-apoptotic cascade activates BID, which oligomerizes BAK or BAX into pores that result in the release of cytochrome cCell Death Differ7200011661173
- Y.LiS.LiX.QinW.HouH.DongL.Yaoet al.The pleiotropic roles of sphingolipid signaling in autophagyCell Death Dis520141245
- M.A.LevyH.FutermanMammalian ceramide synthasesIUBMB Life622010347356
- C.R.GaultL.M.ObeidY.A.HannunAn overview of sphingolipid metabolism: from synthesis to breakdownAdv Exp Med Biol6882010123
- Y.A.C.LubertoK.M.ArgravesEnzymes of sphingolipid metabolism: from modular to integrative signalingBiochemistry40200148934903
- S.A.F.MoradJ.C.LevinS.F.TanT.E.FoxD.J.FeithM.C.CabotNovel off-target effect of tamoxifen - inhibition of acid ceramidase activity in cancer cellsBiochim Biophys Acta1831201316571664
- R.KolesnickThe therapeutic potential of modulating the ceramide/sphingomyelin pathwayJ Clin Invest1200238
- S.Y.V.ChanA.L.HilchieM.G.BrownR.AndersonD.W.HoskinApoptosis induced by intracellular ceramide accumulation in MDA-MB-435 breast carcinoma cells is dependent on the generation of reactive oxygen speciesExp Mol Pathol822007111
- A.BaiC.MaoR.W.JenkinsZ.M.SzulcA.BielawskaY.A.HannunAnticancer actions of lysosomally targeted inhibitor, LCL521, of acid ceramidasePLoS One122017 e0177805
- N.C.ZitomerT.MitchellK.A.VossG.S.BondyS.T.Pruettet al.Ceramide synthase inhibition by fumonisin B1 causes accumulation of 1-deoxysphinganine: a novel category of bioactive 1-deoxysphingoid bases and 1-deoxydihydroceramides biosynthesized by mammalian cell lines and animalsJ Biol Chem284200947864795
- M.BektasC.E.OrfanosC.C.GeilenDifferent vitamin D analogues induce sphingomyelin hydrolysis and apoptosis in the human keratinocyte cell line HaCaTCell Mol Biol (Noisy-le-grand)462000111119
- A.BettaïebI.PloV.Mansat-De MasA.Quillet-MaryT.Levadeet al.Daunorubicin-and mitoxantrone-triggered phosphatidylcholine hydrolysis: implication in drug-induced ceramide generation and apoptosisMol Pharmacol551999118125
- N.BeckmannD.SharmaE.GulbinsK.A.BeckerB.EdelmannInhibition of acid sphingomyelinase by tricyclic antidepressants and analogonsFront Physiol52014331
- M.Mashhadi Akbar BoojarM.Mashhadi Akbar BoojarS.GolmohammadI.BahrehbarData on cell survival, apoptosis, ceramide metabolism and oxidative stress in A-494 renal cell carcinoma cell line treated with hesperetin and hesperetin-7-O-acetateData in Brief202018596601
- V.Gouazé-AnderssonM.FlowersR.KarimiG.FabriásA.Delgadoet al.Inhibition of acid ceramidase by a 2-substituted aminoethanol amide synergistically sensitizes prostate cancer cells to N-(4-hydroxyphenyl) retinamideProstate71201110641073
- S.P.SardiC.VielJ.ClarkeC.M.TreleavenA.M.Richardset al.Glucosylceramide synthase inhibition alleviates aberrations in synucleinopathy modelsProc Natl Acad Sci USA114201726992704
- M.Mashhadi Akbar BoojarM.HassanipourS.Ejtemaei MehrM.Mashhadi Akbar BoojarA.R.DehpourNew aspects of Silibinin stereoisomers and their 3-O-galloyl derivatives on cytotoxicity and ceramide metabolism in Hep G2 hepatocarcinoma cell lineIran J Pharm Res152016421433
- F.M.GonA.AlonsoSphingomyelinases: enzymology and membrane activityFEBS Lett53120023846
- N.Andrieu-AbadieT.LevadeSphingomyelin hydrolysis during apoptosisBiochim Biophys Acta15852002126134
- N.MarchesiniY.A.HannunAcid and neutral sphingomyelinases: roles and mechanisms of regulationBiochem Cell Biol8220042744
- J.H.ParkE.H.SchuchmanAcid ceramidase and human diseaseBiochim Biophys Acta1758200621332138
- X.LiuJ.C.ChengL.S.TurnerS.ElojeimyT.H.BeckhamA.Bielawskaet al.Acid ceramidase up-regulation in prostate cancer: role in tumor development and implications for therapyExpert Opin Ther Targets13200914491458
- M.Mashhadi Akbar BoojarM.Mashhadi Akbar BoojarS.GolmohammadM.NikkhahYazdiCeramide generation as a novel biological mechanism for chemo-preventive and cytotoxic effects of hesperidin on HT-144 melanoma cells. Beni-Suef UniversityJ Basic Appl Sci2018 IN Press
- D.ProkschJ.J.KleinC.ArenzPotent inhibition of acid ceramidase by novel B-13 analoguesJ Lipids201120118
- Y.Y.LiuY.T.LiCeramide glycosylation catalyzed by glucosylceramide synthase and cancer drug resistanceAdv Cancer Res11720145989
- J.F.LiS.J.ZhengL.L.WangS.LiuF.Renet al.Glucosylceramide synthase regulates the proliferation and apoptosis of liver cells in vitro by Bcl-2/Bax pathwayMol Med Rep16201773557360
- M.KartalG.SaydamF.SahinY.BaranResveratrol triggers apoptosis through regulating ceramide metabolizing genes in human K562 chronic myeloid leukemia cellsNutr Cancer632011637644
- A.Gomez-MuñozThe role of ceramide 1-phosphate in tumor cell survival and disseminationAdv Cancer Res1402018217234
- K.KitataniJ.Idkowiak-BaldysJ.BielawskiR.W.JenkinsC.E.Senkalet al.Protein kinase C-induced activation of a ceramide/protein phosphatase 1 pathway leading to dephosphorylation of p38 MAPKJ Biol Chem281320063679336802
- R.KolesnickZ.FuksRadiation and ceramide-induced apoptosisOncogene22200358975906
- S.LahiriA.H.FutermanThe metabolism and function of sphingolipids and glycosphingolipidsCell Mol Life Sci64200722702284
- K.PalmanoA.RowanR.GuillermoJ.GuanP.M.JarrowThe role of gangliosides in neurodevelopmentNutrients7201538913913
- J.MatsudaM.T.VanierI.PopaJ.PortoukalianK.SuzukiGD3- and O-acetylated GD3-gangliosides in the GM2 synthase-deficient mouse brain and their immunohistochemical localizationProc Jpn Acad Ser B Phys Biol Sci822006189196
- P.HerseyO.JamalC.HendersonI.ZardawiG.D'AlessandroExpression of the gangliosides GM3, GD3 and GD2 in tissue sections of normal skin, naevi, primary and metastatic melanomaInt J Cancer411988336343
- S.CaughlinS.MaheshwariN.WeishauptK.K.YeungD.F.CechettoS.N.WhiteheadAge-dependent and regional heterogeneity in the long-chain base of A-series gangliosides observed in the rat brain using MALDI ImagingSci Rep7201716135
- R.B.ChanA.J.PerotteB.ZhouC.LiongE.J.ShorrK.S.Marderet al.Elevated GM3 plasma concentration in idiopathic Parkinson's disease: a lipidomic analysisPLoS One122017 e0172348
- A.Di PardoV.MaglioneSphingolipid metabolism: a new therapeutic opportunity for brain degenerative disordersFront Neurosci17122018249
- Y.IlanCompounds of the sphingomyelin-ceramide-glycosphingolipid pathways as secondary messenger molecules: new targets for novel therapies for fatty liver disease and insulin resistanceAm J Physiol Gastrointest Liver Physiol3102016G1102G1117
- B.OgretmenSphingolipid metabolism in cancer signalling and therapyNat Rev Cancer1820183350
- M.C.PetersenG.I.ShulmanRoles of diacylglycerols and ceramides in hepatic insulin resistanceTrends Pharmacol Sci382017649665